Russianthistle
Grasshopper
|
|
|
|
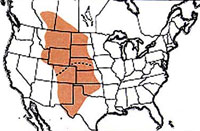
Geographic range of Aeoloplides turnbulli (Thomas) |
|
|
Distribution
and Habitat
Economic
Importance
Food
Habits
Migration
and Dispersal
Identification
Hatching
Nymphal
Development
Adults
and Reproduction
Population
Ecology
Daily
Activity
Selected
References
Click here for the printable version
Distribution and Habitat
The Russianthistle grasshopper enjoys a wide geographic range in western North America from southern Canada to northern Mexico. It inhabits desert shrub sites, local alkali flats vegetated with saltbush and black greasewood, and grasslands, where it does not feed on grasses but relies on the presence of host forbs, species of the goosefoot family (Chenopodiaceae).
Economic Importance
Depending on the vegetation of its habitat or the kind of crop growing nearby, the Russianthistle grasshopper is either a beneficial or a pest species. In western Kansas in 1938, outbreak populations caused severe damage to sugar beets. Two years later during the same outbreak, numerous individuals invaded wheat fields in late June but caused no damage to the crop because of an abundant supply of green Russianthistle, a preferred host plant.
Populations that irrupt in desert shrub communities, such as the Big Horn Basin, Wyoming, damage valuable forage chenopods including Gardner saltbush, fourwing saltbush, and winterfat. Livestock, especially sheep, select these shrubs for feed during winter months when grasses and forbs are scarce or absent.
This grasshopper plays a beneficial role when populations inhabit patches of kochia, Russian thistle, or lambsquarters growing on disturbed land and in field borders and roadsides. The grasshoppers feed on the growing points of these plants, reducing seed production and foliage.
The Russianthistle grasshopper is in the largest third of rangeland grasshoppers. Live weight of males and females collected from a desert shrub community in the Big Horn Basin, Wyoming, with Gardner saltbush the chief host plant, averaged 296 mg and 548 mg, respectively (dry weights: 94 mg and 175 mg, respectively). Live weight of males and females collected from a disturbed site in southeast Wyoming invaded by kochia averaged 242 mg and 462 mg, respectively (dry weights: 76 mg and 146 mg).
Food Habits
The Russianthistle grasshopper feeds on species of the goosefoot family of plants (Chenopodiaceae). Principal host plants are shrubs or stout herbs, which afford the grasshoppers both nutritious food and roosting sites. Shrubs that serve commonly as host plants include fourwing saltbush, Gardner saltbush, white sage, also called winterfat, (Ceratoides lanata), and black greasewood, while stout herbs include the weeds kochia, Russian thistle, and lambsquarters. The latter plants frequently occur together in disturbed land providing the grasshoppers with an opportunity for a mixed diet. Likewise, in desert shrub habitats several chenopod species are usually present in addition to the principal host plant. In a desert shrub site of the Big Horn Basin, Wyoming, Gardner saltbush was the principal host plant but other chenopods were available to the grasshoppers. These were chiefly small annuals with five species sustaining grasshopper damage: Atriplex argenta, A. powelli, A. suckleyi, Sarcobatus vermiculatus, and Suaeda nigra. One species showed no damage, the introduced Halogeton glomeratus. This is unfortunate because this weed is very difficult to control and is toxic to sheep due to its high content of oxalates.
Field observations and gut analyses indicate that the Russianthistle grasshopper also feeds sparingly on several forbs belonging to at least six plant families other than Chenopodiaceae.
The Russianthistle grasshopper's method of attacking its host plant is notable for the selection of young tissue. From basking or roosting positions on kochia or Gardner saltbush, the grasshoppers climb to the growing tips to feed. There they consume young leaves and other developing tissues. Timed feeding bouts of two adults took six and seven minutes. In the desert shrub community of the Big Horn Basin, where Gardner saltbush dominates and much bare soil prevails, both nymphs and adults often come down to the ground during the day. They do not, however, appear to feed on ground litter.
Dispersal and Migration
Very little is known about the dispersal and migration of the Russianthistle grasshopper. Two accidentals, a male and a female of the southern subspecies, Aeoloplides turnbulli bruneri, were found in the mountains west of Boulder, Colorado. One was an adult male taken at 7,700 feet on 7 August 1933, the other an adult female taken at 12,100 feet on 4 August 1960. Collection of these two provides evidence for dispersal of the long-winged subspecies, A. t. bruneri.
Wings of the northern subspecies, A. t. turnbulli, vary in length but are usually noticeably short of the end of the abdomen. Compare Figure 7 of a female of the northern subspecies collected in eastern Wyoming (Platte County) with Figure 8 of a female of the southern subspecies collected in southeastern Colorado (Bent County). The wings of adults of the northern subspecies that inhabit the Big Horn Basin, Wyoming (Big Horn County) are even shorter, falling short of the end of the hind femur by 3 to 6 mm. The hind wings of the males averaged 8.8 mm long and of females 9.7 mm. In spite of the relatively short wings and heavy weight, the adults are able to fly. In flushed flight they travel an average distance of 3.6 feet at heights of 4 to 6 inches. The flight is silent and straight with the grasshopper usually landing face away from the intruder.
Identification
|
Figures 1-5. Appearance
of the nymphal instars, their sizes, structures, and color
patterns. Notice progressive development of wing pads. BL=body
length, FL=Hind Femur Length. AS=number of antennal segments
|
 |
|
Fig. 1,
First Instar: BL 3.8-4.9 mm FL 2.5-2.8 mm AS 12-14.
|
 |
|
Fig. 2,
Second Instar: BL 5.6-7.1 mm FL 3.6-4.3 mm AS 15-17.
|
 |
|
Fig. 3,
Third Instar: BL 7.7-8.5 mm FL 5.0-5.9 mm AS 18-20.
|
 |
|
Fig. 4,
Fourth Instar: BL 10-13 mm FL 6.5-7.1 mm AS 20-22.
|
 |
|
Fig. 5,
Fifth Instar: BL 14-17.3 mm FL 9-10.3 mm AS 22-24.
|
|
Figures 6-10. Appearance
of the adult male and 2 females, cercus, and egg pod.
|
 |
|
Fig. 6,
Adult Male: BL 18-20.5 mm FL 10-11.2 mm AS
23-26. Northern subspecies A. turnbulli turnbulli
|
 |
|
Fig. 7,
Adult Female: BL 18.8-25
mm FL 11-13 mm AS 23-25. Northern subspecies A. turnbulli
turnbulli
|
 |
|
Fig. 8, BL 21-23.5
mm FL 11-12.5 mm AS 22-24. Southern subspecies A. turnbulli
bruneri
|
 |
|
Fig. 9, End of male
abdomen showing cercus and subgenital plate with subapical
tubercle.
|
 |
|
Fig. 10, Egg pod and
exposed eggs in opened pod.
|
The Russianthistle grasshopper is a medium-sized robust species. It has a distinctive color pattern of shades of tan with fuscous markings (Figs. 6 and 7). The wings of the northern subspecies, A. t. turnbulli, may be 2 to 4 mm short of the end of the abdomen (and also the end of the hind femur, a less variable measurement). The wings of the southern subspecies, A. t. bruneri, are longer, surpassing the end of the abdomen (Fig. 8). Lateral light bands mark the sides of the pronotal disk and are separated by a broad fuscous band. The latter band is divided by the median carina, which may be colored partly or entirely light tan. The hind femur has three characteristic fuscous marks on the medial area. These dark marks usually extend dorsally onto the upper marginal areas. The hind tibia is usually medium blue with a pale tan annulus near the base. The male possesses tiny furcula represented by minute lobes, a subgenital plate with a conspicuous subapical tubercle, and triangular slender cerci (Fig 9).
The nymphs ( Fig. 1-5)are identifiable by their structure, color patterns, and shape.
- Head with face nearly vertical, head tan to pale tan or green with several to many fuscous spots; compound eye with large pale tan spots in brown reticulum; antennae filiform, first two segments (scape and pedicel) pale yellow with several brown spots, rest of segments (flagellum) fuscous, each with pale anterior annulus.
- Pronotum tan or green with several to many brown spots, median carina pale contrasting with main color of disk, the pale line extends onto abdomen; disk becomes distinctly wider posteriorly (a diagnostic character).
- Outer medial area of hind femur uniformly tan or green with several to many brown spots or with three fuscous markings, tan, and spotted (Fig. 2). Hind tibia is green, blue or pale gray, usually with brown spots.
- General body color tan or green.
Characters especially useful in identifying the nymphs are: 1. the shape of the pronotal disk, which widens from front to rear, 2. most individuals heavily spotted brown or fuscous, 3. nymphs are colored pale green, pale tan, or pale gray, 4. in tan or gray nymphs, three characteristically shaped dark markings are usually present on the medial and marginal areas of the hind femur (Fig. 1 and 2), in green forms, these markings are faint or lacking (Fig. 3).
Hatching
The Russianthistle grasshopper is an early-hatching species. In weedy roadside habitats of eastern Wyoming, first instars appear as early as late April or the first of May along with the twostriped grasshopper, Melanoplus bivittatus. In the desert shrub of the Big Horn Basin, hatching may occur one to two weeks earlier. The hatching period is extended, lasting from four to five weeks. No study of egg development has been made.
Nymphal Development
Both the male and female nymphs require five instars to develop to the adult stage. Because of their nature to develop a robust body, their early hatching in spring, and their habitual location high on the host plant where ambient temperatures are lower than on the ground, the Russianthistle grasshopper's nymphal period is relatively prolonged ranging from 50 to 60 days.
Adults and Reproduction
In the desert shrub habitat of the Big Horn Basin, adults may appear as early as mid June, but in the weedy roadsides and disturbed sites of southeastern Wyoming, adults of the Russianthistle grasshopper do not appear until the last week of June. The processes of sexual maturation and courtship have not been determined in this species. The duration of copulation by a pair is unknown, but mating pairs have been observed sitting on host plants and on the ground during July and August from early morning until late evening. Attempts at oviposition by two females were observed in the desert shrub habitat of the Big Horn Basin. The first observation was made on 17 August 1993 from 1 to 1:07 p.m. DST when soil surface temperature was 120°F, air temperature 82°F, and sky clear. The female was discovered walking and hopping on the ground. After traveling a distance of 12 feet, she stopped and began to bore into bare ground for several seconds, then withdrew her ovipositor and walked another 12 feet. When she stopped again at 1:05 p.m., she bore into the soil for approximately one minute. She again withdrew her ovipositor and walked. By this time she was some distance from the observer and was lost to view. In the same desert shrub site another female was observed attempting to oviposit on 19 August 1993 at 1:23 p.m. DST when soil surface temperature was 121°F, air temperature 83°F, and sky clear. After this female was discovered, she crawled 2 feet into the shade of a clump of halogeton. She began to bore into the soil at 1:23 p.m. and continued for four minutes until she was disturbed by the observer and walked away, disappearing into the vegetation. These two attempts at oviposition suggest that the Russianthistle grasshopper oviposits in bare ground. The pods are stubby, 7/16 inch long, 3/16 inch diameter, and slightly curved. They have a sloping cap and the top appears pinched (Fig. 10). A plug of froth is lacking but reddish brown froth completely surrounds the egg mass and the individual eggs. The eggs are light-reddish brown or tan and 4.0 to 4.4 mm long. There are 12 to 18 eggs in a pod. The fecundity of females is high. In Canada, six caged females produced an average of 250 eggs each.
Population Ecology
Populations of the Russianthistle grasshopper develop in sites with an abundant supply of host plants. Such habitats usually occur in disturbed land, crop borders, weedy rangeland, and desert shrub associations dominated by saltbush (Atriplex spp.). The high fecundity of this species allows populations to irrupt during environmentally favorable periods. Densities in farmyard patches of weeds, usually less than an acre in size, may reach 100 or more young adults per square yard.
Outbreaks may occur in much larger areas such as the wheat-growing region of western Kansas and eastern Colorado. During the drought period of 1931-41, A. t. bruneri came into prominence in 1937. The grasshoppers hatched along the edges of wheat fields where Russian thistles had grown profusely the previous fall. The species became the dominant grasshopper in this area from 1938 to 1941 supplanting the migratory grasshopper, Melanoplus sanguinipes, as the most abundant species. In a 10 square mile study area of Finney County, Kansas, the Russianthistle grasshopper averaged one young adult per square yard in 1939. In 1940 the population more than doubled, and in 1941 the population remained high early in the season. During the latter half of May 1941, 150 late instar nymphs per square yard were recorded. But in June an epizootic struck the population and brought the outbreak to an abrupt end. Caused by an undetermined species-specific pathogen, the disease almost completely obliterated the Russianthistle grasshopper from the entire area.
Outbreaks of the northern subspecies, A. t. turnbulli, also occur. In 1993 an outbreak of this grasshopper was discovered in a desert shrub habitat of the Big Horn Basin, Wyoming. The vegetation of the infested area, at least 10 square miles in extent, was dominated by Gardner saltbush, a preferred host plant. High densities of this grasshopper ranging from 8 to 13 young adults per square yard occurred everywhere within the area. The adult population sustained a low daily mortality rate of 5 percent. The outbreak has continued for a minimum of two years, as high numbers were present in 1994.
Despite the prevalence of a variety of host plants (chenopods), few rangeland sites of the mixedgrass prairie harbor populations of the Russianthistle grasshopper. Only four sites out of 241 (1.7 percent) surveyed in eastern Wyoming in 1991 and only two sites out of 42 (1 percent) surveyed in the mixedgrass prairie of Montana in 1953 and 1954 contained this species. However, in 1991 in the desert shrub of the Big Horn Basin, this species occupied 23 sites out of 190 (12 percent) surveyed. Densities of grasshoppers in an eastern Wyoming rangeland site with an abundance of host plants of the Russianthistle grasshopper (Russian thistle, winterfat, and slimleaf lambsquarters) were sampled for several years. This survey revealed that for an eight-year period the populations of Russianthistle grasshoppers fluctuated from less than 0.1 per square yard to 0.54 per square yard and that the species held a low rank of abundance in the grasshopper assemblage in all years.
Daily Activity
The Russianthistle grasshopper is a phytophilous species spending much time on its host plant. However, during daylight hours in habitats of sparse vegetation with an abundance of bare soil, the grasshoppers descend regularly to the ground. There they bask, rest, crawl, and disperse short distances.
Extended observations of the species' behavior were made in a native desert shrub habitat of the Big Horn Basin, Wyoming, where Gardner saltbush served as its principal host plant. During the night nymphs and adults usually rest vertically, head-up on the stems of this shrub; a few take positions on ground litter underneath the shrub. One hour after sunrise the grasshoppers resting on the host plant begin to bask by adjusting their positions so that the rays of the sun strike a side or their back. They may also lower a hindleg to expose the abdomen more fully. A few individuals may jump to the ground to bask. Basking lasts for about two hours, after which the grasshoppers become active. Some may crawl to the top of the plant to feed on the growing tips, some may undergo a molt, some may jump from one stem to another, and many continue to roost quietly. Eventually the majority leave the host plant and spend time on the ground during the day until evening when they again seek shelter of the host plant. On the ground they mix among themselves and disperse short distances. Many mating pairs occupy both the host plant and the ground surface. During five-hour periods of observation, 37 single adults and 12 mating pairs were noted on 21 July 1993 and 17 singles and 13 mating pairs on 17 August 1993. Only one attempt at mating was observed; a female resting on saltbush was mounted by a male on 17 August 1993 at 1:54 p.m. The encounter ended unsuccessfully with no consummation of mating. In early afternoon females probed the soil for oviposition but no actual laying of eggs was observed.
When soil surface temperatures are above 120°F during midday, grasshoppers on the ground take evasive actions to prevent overheating. They may directly face away from the sun and stilt, or they may crawl and hop into the shade of vegetation or under the canopy of saltbush. Because the grasshoppers located on the host plant are not subjected to the high ground temperatures, they continue their activities and roosting.
When temperatures cool in the afternoon the grasshoppers on the ground become active again. Continued cooling of the habitat incites the grasshoppers both on the ground and on the host plant to assume basking orientations. Finally, close to sunset, the grasshoppers seek shelter. Those on bare ground crawl to the saltbush host plant; some rest on litter under the bush and many crawl up a stem and rest within the canopy.
In dense stands of kochia and lambsquarters the behavior of the Russianthistle grasshopper differs somewhat from that of the grasshoppers occupying habitats with much bare ground. In dense vegetation the grasshoppers roost all day, most vertically head-up on plant stems, but some horizontally on leaves. Rarely coming down from their perches, they carry out essential activities on the host plants - basking, feeding, molting, and mating. Gravid females, however, are compelled to descend to bare ground to oviposit into soil.
Selected References
Alexander, G. and J. R. Hilliard, Jr. 1969. Altitudinal and seasonal distribution of Orthoptera in the Rocky Mountains of northern Colorado. Ecol. Monogr. 39: 385-431.
Anderson, N. L. 1973. The vegetation of rangeland sites associated with some grasshopper studies in Montana. Montana Agr. Exp. Stn. Bull. 668.
Newton, R. C. and A. B. Gurney. 1956. Distribution of range grasshoppers. Coop. Econ. Insect Report 6: 600.
Vickery, V. R. and D. K. McE. Kevan. A monograph of the Orthopteroid insects of Canada and adjacent regions. Lyman Entomol. Mus. and Res. Lab. Mem. 13 Vol 2: 740-745.
Wallace, H. S. 1955. Revision of the genus Aeoloplides (Orthoptera, Acrididae). Ann. Entomol. Soc. Am. 48: 453-480.
Wilbur, D. A. and R. F. Fritz. 1942. An epizootic among the thistle hoppers, Aeoloplus turnbulli bruneri. Caud., in Kansas. J. Econ. Entomol. 35: 109.