Eleutherengonides
Superorder Acariformes
Order Trombidiformes
Suborder Prostigmata
Supercohort Eleutherengonides
Cohort (Superfamily): Raphignathina (Raphignathoidea, Cheyletoidea, Tetranychoidea, Pterygosomatoidea), Heterostigmatina (Tarsocheyloidea, Heterocheyloidea, Pyemotoidea, Pygmephoroidea, Tarsonemoidea)
Common names: spider mites, peacock mites, false spider mites, raphignathoids, cheyletoids, myobiids, cheyletiella, follicle mites, quill mites, skin mites, walking dandruff, tarsonemids, pygmephorids, scutacarids, pyemotids, straw itch mites, red pepper mites, tracheal mites, broad mite, cyclamen mite.
Probability of Encounter: Very high
Quarantine importance: Very high. The Tetranychoidea contains most of the important plant-parasitic mites outside of the Eupodides, including the spider mites and false spider mites. The Cheyletoidea contains many important parasites of wildlife and some important pests of pets, laboratory animals, livestock, and people. The Heterostigmatina contains a couple of important crop pests (broad mite, cyclamen mite) and some pests of ferns, mushroom culture (red pepper mites), and bees (tracheal mites), as well as parasitoids of insects, including those that sometimes bite people (straw itch mites).
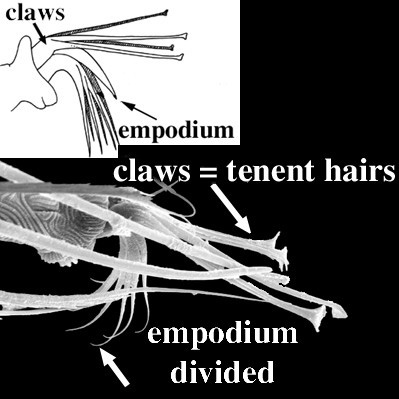
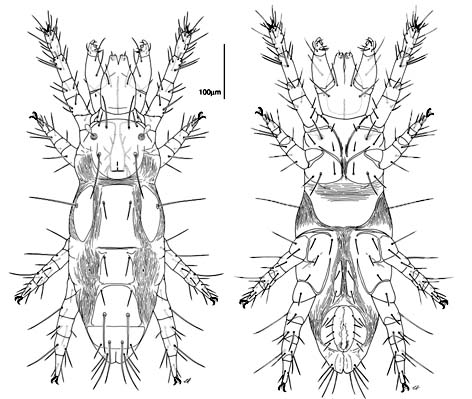
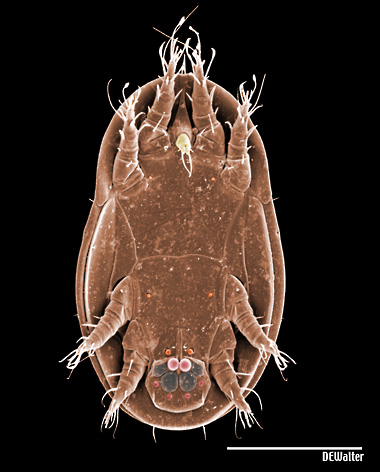
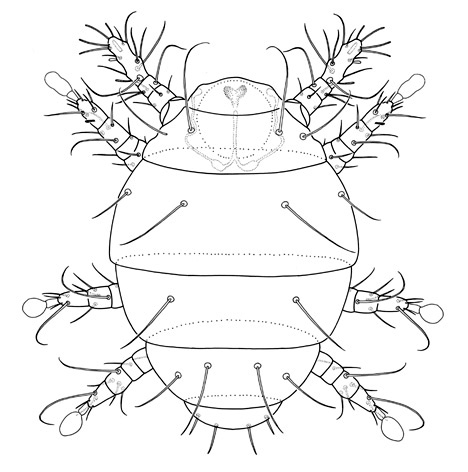
Diagnosis. Raphignathina are small to medium in size; most species soft-bodied, others with varying degrees of dorsal sclerotization; white, red, yellow, green or brown in colour. Cheliceral bases adnate, fused mesally, or fused with the subcapitulum; chelicerae with fixed digit reduced and movable digit bladelike, needle-like, or whiplike. Peritremes, when present, often elaborate chambered structures on the dorsal surface of the chelicerae; naso absent; prodorsal trichobothria absent; eye lenses usually present. Palps 1-5 segmented; usually usually with one or more claw-like setae on the palp tibia and with a thumb- or button-like subterminal palp tarsus. Body setae range from short and sparse, to long and barbed, to dentritic, to plate-like. Leg tarsi usually with tenent hairs on claws or empodium. Genital papillae absent; males often with an intromittent aedeagus.
Heterostigmatina are minute to
medium-sized; usually well armoured, with several large dorsal
sclerites; white, yellow, or brown in
colour. Capitulum
typically head-like; cheliceral bases fused into a
single unit (stylophore) that may or may not be fused with the subcapitulum;
fixed digit of the chelicera absent, movable digit blade-like or
styletiform. Palps linear and reduced
to 3 or fewer segments; palps may appear to be absent. In females stigmata open anterolaterally on
the ‘shoulders’ of the prodorsum, and the single pair of prodorsal
trichobothria is capitate; stigmata and trichobothria absent in males (except
Tarsocheyloidea); eyes absent.
Dorsal setation reduced. Legs I
often with tarsal claw modified as a hook; legs IV in females often reduced or
absent; coxal fields fused to varying degrees so that apodemes form cross-like
patterns ventrally. Genital papillae
absent. Many species exhibit
polymorphism.
Similar taxa. Anystina
and Eupodina usually have genital papillae and
prodorsal trichobothria, both of which are lacking in the Raphignathina.
Ecology & Distribution. The Raphignathoidea is composed of several families of predatory and herbivorous mites, and are very common in soil, moss and on foliage. The Stigmaeidae are the most ecologically diverse, acting as predators in leaf litter and in trees, and herbivores in moss. Barbutiidae, Caligonellidae, Camerobiidae, Cryptognathidae, Dasythyreidae, Eupalopsellidae and Raphignathidae are frequently encountered in the drier soil microhabitats (e.g. dry soil, bark, moss). Homocaligidae, on the other hand, appear restricted to semi-aquatic habitats. Most families of Cheyletoidea are parasites that are unlikely to turn up in soil samples. The exception is Cheyletidae, which includes many free-living predators inhabiting litter, soil, tree bark, foliage, as well as stored products and vertebrate nests. The Tetranychoidea are plant-associated mites, and includes the infamous spider mites (Tetranychidae). Tetranychoid species that feed on grasses or pasture legumes may occasionally appear in soil samples.
Most
families of Heterostigmata are associated with insects and often turn up in
pitfall traps or leaf litter extractions when their hosts fall into collecting
vials. Tarsocheylids have been
collected from the tunnels of bark beetles, beneath the elytra of passalid
beetles, as well as from rotting wood.
Heterocheylids are also passalid associates. Neither of these taxa is known to feed on their hosts. The interactions of Pygmephoroidea with
their hosts are primarily phoretic, and the mites feed on fungus upon reaching
their destination. In contrast, many
species in the Pyemotoidea are parasites, parasitoids, and predators of
insects. They are often found in
infested stored products, and their mistaken biting of humans can cause severe
itching and allergic reactions (‘hay itch’, ‘straw itch’, and ‘grocer’s
itch’). Female morphs specialised for
hanging on to their hosts – phoretomorphs
occur in several families of pygmephoroids and pyemotoids. These polymorphisms have caused much
taxonomic confusion within the Heterostigmata.
Within the Tarsonemoidea, the Podapolipidae are parasites of Coleoptera,
Orthoptera and Hymenoptera. The Tarsonemidae
have the most varied ecology of the Heterostigmata, sometimes acting as parasites
or predators of insects (the most famous of these being the honeybee parasite
Acarapis woodi), sometimes as fungivores, and sometimes as plant
parasites. Numerous economically
important pests fall into this last group.
References
Clift, AD. & Toffolon RB. 1981.
Biology, fungal host preferences and economic significance of two pygmephorid
mites (Acarina : Pygmephoridae) in cultivated mushrooms, N.S.W., Australia. Mushroom
Science 11: 245-253.
Evans GO. 1992. Principles of
Acarology. CABI, Wallingford.
Husband RW. 1990. New species of Podapolipoides
(Acari : Podapolipidae), ectoparasites of grasshoppers (Orthoptera : Acrididae)
in Australia and New Zealand, with keys to world species. Ann. Ent. Soc. Amer. 83 : 371-393.
Kethley JB.
1982. Acariformes. In: Parker, S.P. (ed.) Synopsis and Classification of Living
Organisms. McGraw-Hill, New York,
pp. 142-145.
Kethley JB.
1990. Acarina: Prostigmata
(Actinedida). In DL Dindal (ed.) Soil Biology Guide. John Wiley & Sons, New York, pp. 667-756.
Krantz GW. 1978. A Manual of Acarology. OSU Bookstores: Corvallis.
Lindquist EE. 1976. Transfer of the
Tarsocheylidae to the Heterostigmata, and reassignment of the Tarsonemina and
Heterostigmata to lower hierarchic status in the Prostigmata (Acari). Can.
Entomologist 108: 23-48.
Lindquist EE. 1986. The world genera
of Tarsonemidae (Acari : Heterostigmata) : A morphological, phylogenetic, and
systematic revision, with a reclassification of family-group taxa in the
Heterostigmata. Mem. Ent. Soc. Can. 136: 1-517.
Luxton M. 1973. Mites of the genus Cryptognathus from Australia, New
Zealand, and Niue Island. Acarologia 15: 53-75.
Mahunka S. 1967. A survey of the scutacarid (Acari :
Tarsonemini) fauna of Australia. Aust. J. Zool. 15: 1299-1323.
Moser JC. 1975. Biosystematics of
the straw itch mite with special reference to nomenclature and dermatology. Trans.
Roy. Ent. Soc., Lond. 127: 185-191.
Moser JC & Cross EA. 1975.
Phoretomorph: A new phoretic phase unique to the Pyemotidae (Acarina :
Tarsonemoidea). Ann. Ent. Soc. Amer 68: 820-822.
Noble JC. et al. 1996a. Soil and litter
microarthropod populations from two contrasting ecosystems in semi-arid eastern
Australia. J. Arid Environ. 32: 329-346.
Noble JC et al. 1996b. Fire studies in mallee (Eucalyptus
spp.) communities of western New South Wales: spatial and temporal fluxes in
soil chemistry and soil biology following prescribed fire. Pacific Cons.
Biol. 2: 398-413.
Wood TG. 1967. New Zealand mites of the family
Stigmaeidae (Acari, Prostigmata). Trans. Roy. Soc. NZ 9: 93-139.
Wood TG. 1969. The Homocaligidae, a new family of
mites (Acari: Raphignathoidea), including a description of a new species from
Malaya and the British Solomon Islands. Acarologia 11: 711-729.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}