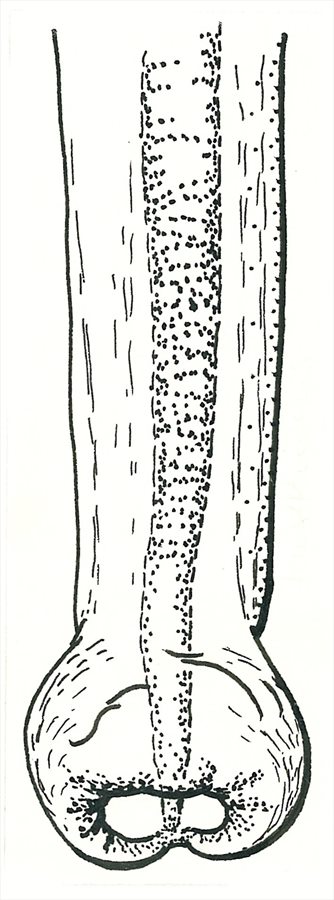
Both sexes fully winged. Body and legs brown to dark brown, fore tarsi light brown, antennal segments III–IV brown but yellow in basal third, V–VI light brown with base paler; fore wing pale with base shaded; major setae pale. Antennae 8-segmented, segment III with 2 sense cones, IV with 4 sense cones; VIII short and broad at base. Head longer than wide; maxillary stylets about one third of head width apart, retracted to compound eyes, maxillary bridge present; postocular setae pointed, about 0.6 as long as dorsal length of compound eye. Pronotum with 5 pairs of major setae with apices pointed, but anteromarginal setae shorter than anteroangulars and neither pair as long as the epimeral and posteroangular pairs; epimeral sutures complete; prosternal basantra present, mesopresternum complete but slender medially. Fore tarsal tooth small. Fore wing constricted medially, with about 10 duplicated cilia; sub-basal setae short and pointed. Tergite IX setae S1 pointed, more than 0.5 as long as tube.
Male with no pore plate on sternite VIII; fore tarsal tooth present; tergite IX setae S2 short and stout; aedeagus with apex slightly swollen.
The genus Haplothrips is one of the three most species-rich genera of Thysanoptera, and currently includes about 245 species worldwide. Most of these species come from the Holarctic or the Old World tropics, with 80 listed from Europe and 14 from Britain. No Haplothrips species is known to be endemic to the Neotropics, although a few are native to southern South America (Mound & Zapater, 2003). Haplothrips species are largely phytophagous, particularly associated with the flowers of Asteraceae and Poaceae, but some are predatory (Mound & Minaei, 2007). Haplothrips hukkineni shares many character states with juncorum, but has the maxillary stylets wider apart medially in the head, and has a shorter tube; furthermore, the aedeagus in males is slightly swollen at the apex rather than lanceolate as in juncorum (Collins, 2012).
Breeding in damp places on the leaves and possibly the inflorescences of its host plants, Bolboschoenus and Cyperus [Cyperaceae], Juncus [Juncaceae], Oryza and Phragmites [Poaceae], also Typha [Typhaceae].
In Britain, collected infrequently and only in the southeast of England, although three times recently on the north Kent coast (Collins, 2012; Badmin, 2016). In Britain, it has only been taken from Bolboschoenus maritimus, and always at the same time as the more common H. juncorum. It is recorded predominantly in central and eastern Europe and the eastern Mediterranean, but also from Norway and Sweden. H. hukkineni has not been recorded from Ireland, but it was erroneously included in the Irish checklist by O’Connor (2008).
PHLAEOTHRIPIDAE - PHLAEOTHRIPINAE
Haplothrips hukkineni Priesner
Haplothrips hukkineni Priesner, 1939: 276
Haplothrips phragmitis Priesner, 1950: 35
Badmin J (2016) Field Meeting Report. Great Bells RSPB reserve, Isle of Sheppey, Kent, 9 August 2015. British Journal of Entomology and Natural History 29: 119–122.
Collins DW (2012) A second British record of Haplothrips hukkineni Priesner (Thysanoptera: Phlaeothripidae). British Journal of Entomology and Natural History 25: 107–108.
Mound LA & Minaei K (2007) Australian insects of the Haplothrips lineage (Thysanoptera – Phlaeothripinae). Journal of Natural History 41: 2919–2978.
Mound LA & Zapater MC (2003) South American Haplothrips species (Thysanoptera, Phlaeothripidae), with a new species of biological control interest to Australia against weedy Heliotropium amplexicaule (Boraginaceae). Neotropical Entomology 32: 437–442.
O’Connor JP (2008) A review of the Irish thrips (Thysanoptera). Irish Naturalists’ Journal 29: 20–24.