
Head of female microptera

Female and male heads
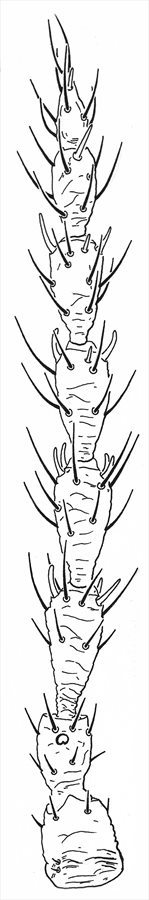
Antenna
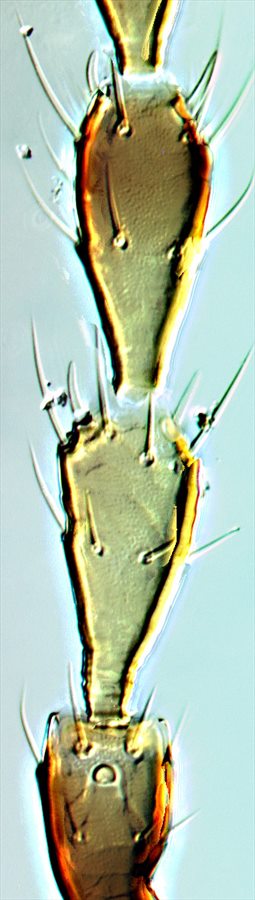
Antennal segments II–IV
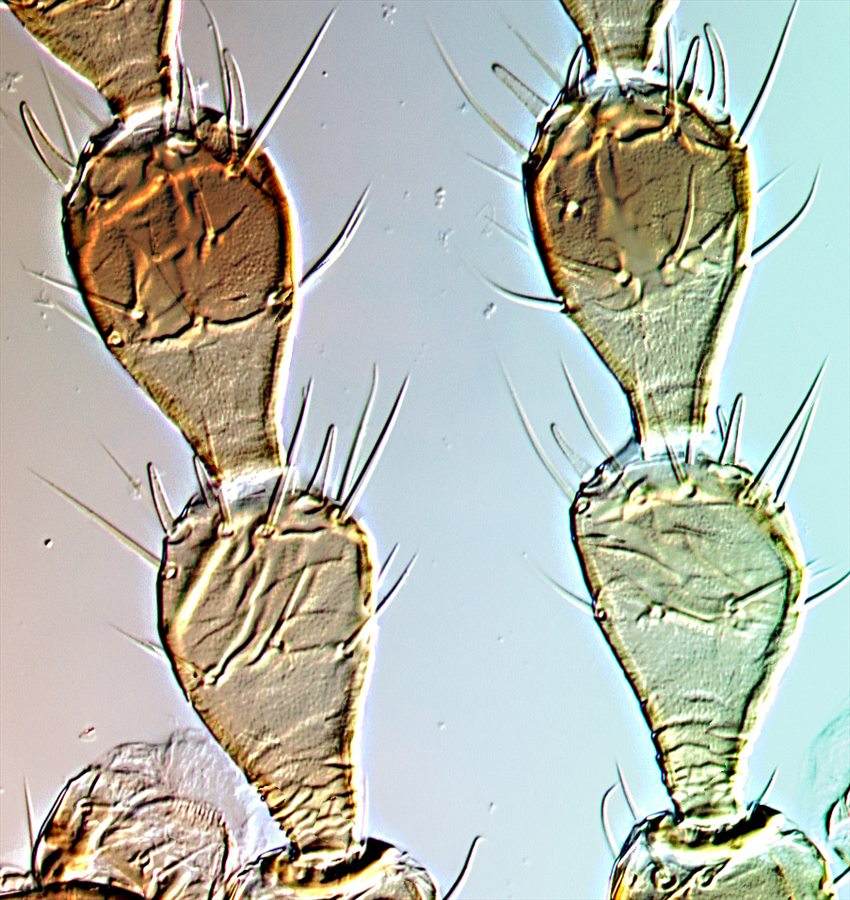
Antennal segments II–IV (distorted in water soluble mountant)
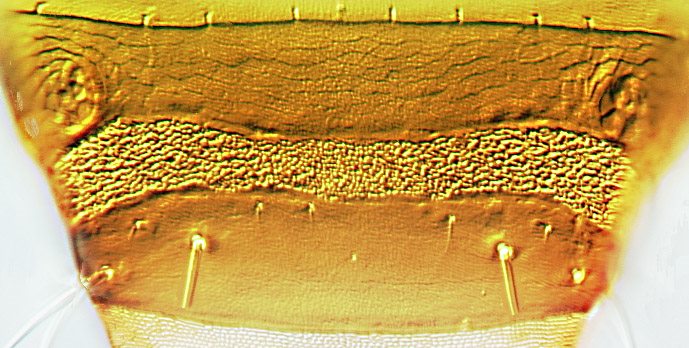
Male sternite VIII
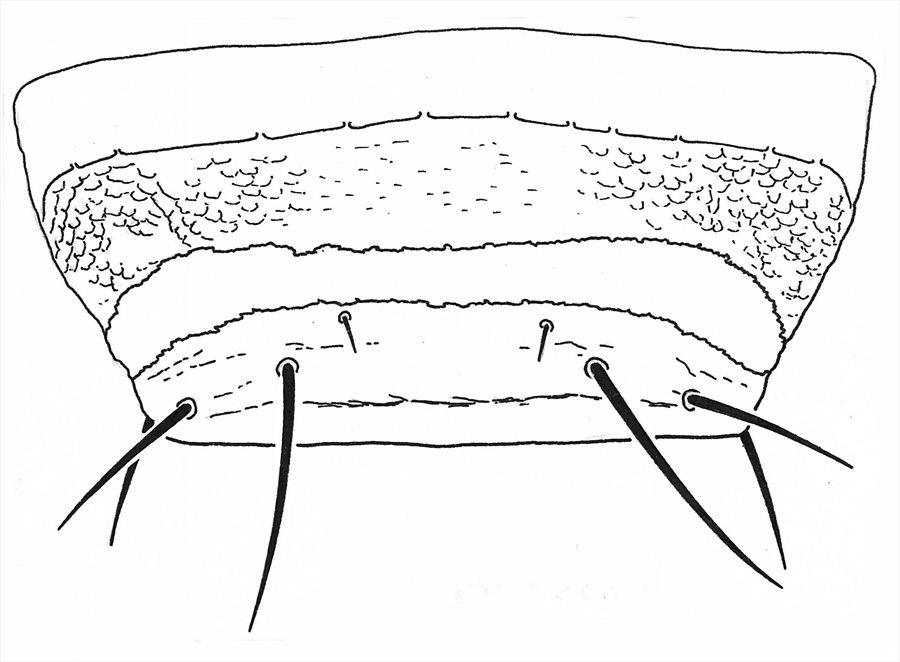
Male sternite VIII
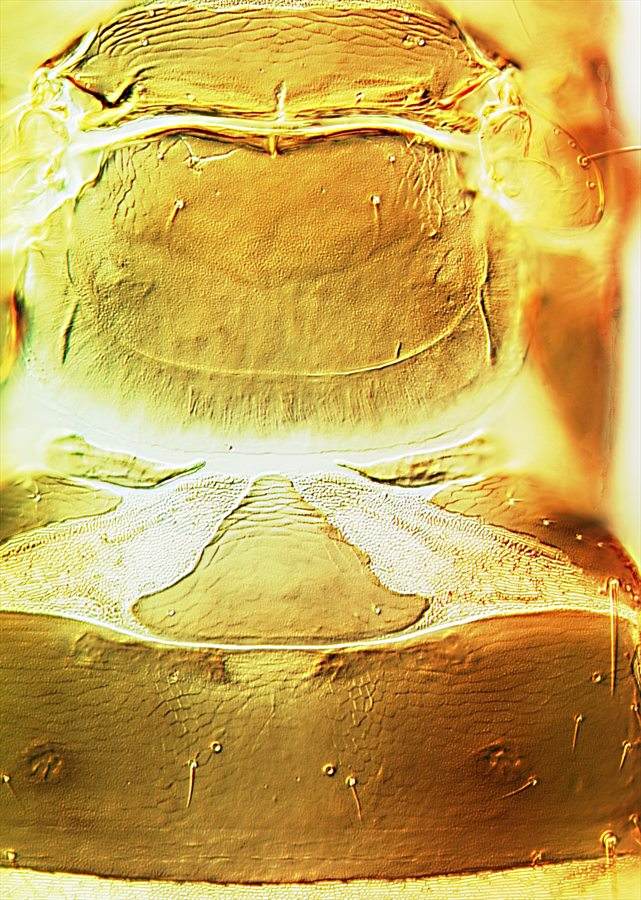
Meso & metanota, pelta & tergite I
Both sexes either fully winged or with wings shorter than thorax width. Body and legs brown, tarsi and much of fore tibiae yellow, mid and hind tibiae yellow at base; antennal segment III mainly yellow, IV–VI variably yellow at base; fore wings weakly shaded toward apex. Antennae 8-segmented; sense cones longer in winged than wingless individuals, segment III with 3 sense cones of which the inner one is short and straight, IV with 4 sense cones; VIII constricted to base. Head longer than wide, wider across cheeks than across eyes; cheeks convex and constricted to base, without prominent tubercles, but with several small, stout setae; postocular setae long and pointed, wide apart; maxillary stylets retracted to eyes, close together medially. Pronotum without sculpture medially, with stout median apodeme in large males; with 4 pairs of slender pointed major setae, anteromarginals no larger than discal setae; prosternal basantra absent. Fore tarsal tooth small in winged, but large in wingless individuals of both sexes. Metanotum without sculpture medially. Fore wing parallel sided, with about 10 duplicated cilia. Abdominal tergite I, the pelta, with lateral margins curving forward away from tergite II; tergites II–VII with two pairs of sigmoid wing-retaining setae, marginal setae S1 long and pointed; tergite IX setae S1 pointed, less than 0.8 as long as tube.
Males varying in size, large males with fore femora swollen; tergite IX setae S2 short and stout; sternite VIII with slender transverse pore plate extending full width of sternite posterior to spiracles, median longitudinal length of pore plate 15–20 microns.
The genus Hoplothrips includes about 120 named species, but there are no modern identification keys to any substantial number of species. The most common species are known to exist as both winged and wingless morphs. Moreover, these species exhibit considerable sexual dimorphism, and males of the same species vary in body size, with some structures exhibiting patterns of allometric growth. As a result, species identification is often difficult (Mound & Walker, 1986; Kobro & Rafoss, 2006; Okajima, 2006). Hoplothrips ulmi is closely similar to H. fungi, but has shorter, straighter, antennal sense cones, and the transverse pore plate on sternite VIII of males is almost parallel-sided with a shorter median longitudinal length. These two species were regarded as synonyms by Schliephake & Klimt (1979), based on specimens from Central Europe. Also related is H. corticis, but that has the posterolateral margins of the pelta confluent with the anterior margin of tergite II, and in males the pore plate on sternite VIII is much larger.
Breeding on dead branches and sometimes forming colonies on the trunk of dead Angiosperm trees, presumably feeding on fungal hyphae.
Widespread and common in England and Scotland, from Kent to northern Scotland (Mound et al., 1976), and recorded widely across central and western Europe, also from Bulgaria, and Japan (Okajima, 2006).
PHLAEOTHRIPIDAE - PHLAEOTHRIPINAE
Hoplothrips ulmi (Fabricius)
Thrips ulmi Fabricius, 1781: 396
Phloeothrips parvipennis Reuter, 1880: 14
Kobro S & Rafoss T (2006) Identification of adult males and females of Hoplothrips species (Thysanoptera: Tubulifera) known from Norway, and some deductions on their life history. Entomologica Fennica 17: 184–192.
Mound LA, Morison GD, Pitkin BR & Palmer JM (1976) Thysanoptera. Handbooks for the Identification of British Insects 1 (11): 1–79.
Mound LA & Walker AK (1986) Tubulifera (Insecta: Thysanoptera). Fauna of New Zealand 10: 1–140.
Okajima S (2006) The Suborder Tubulifera (Thysanoptera). The Insects of Japan 2: 1–720.
Schliephake G & Klimt K (1979) Thysanoptera, Fransenflügler. In: Hannemann, H. J., Schumann, H. & Senglaub, K. (eds): Die Tierwelt Deutschlands 66: 5–477. Jena (G. Fischer).
