Click on images to enlarge
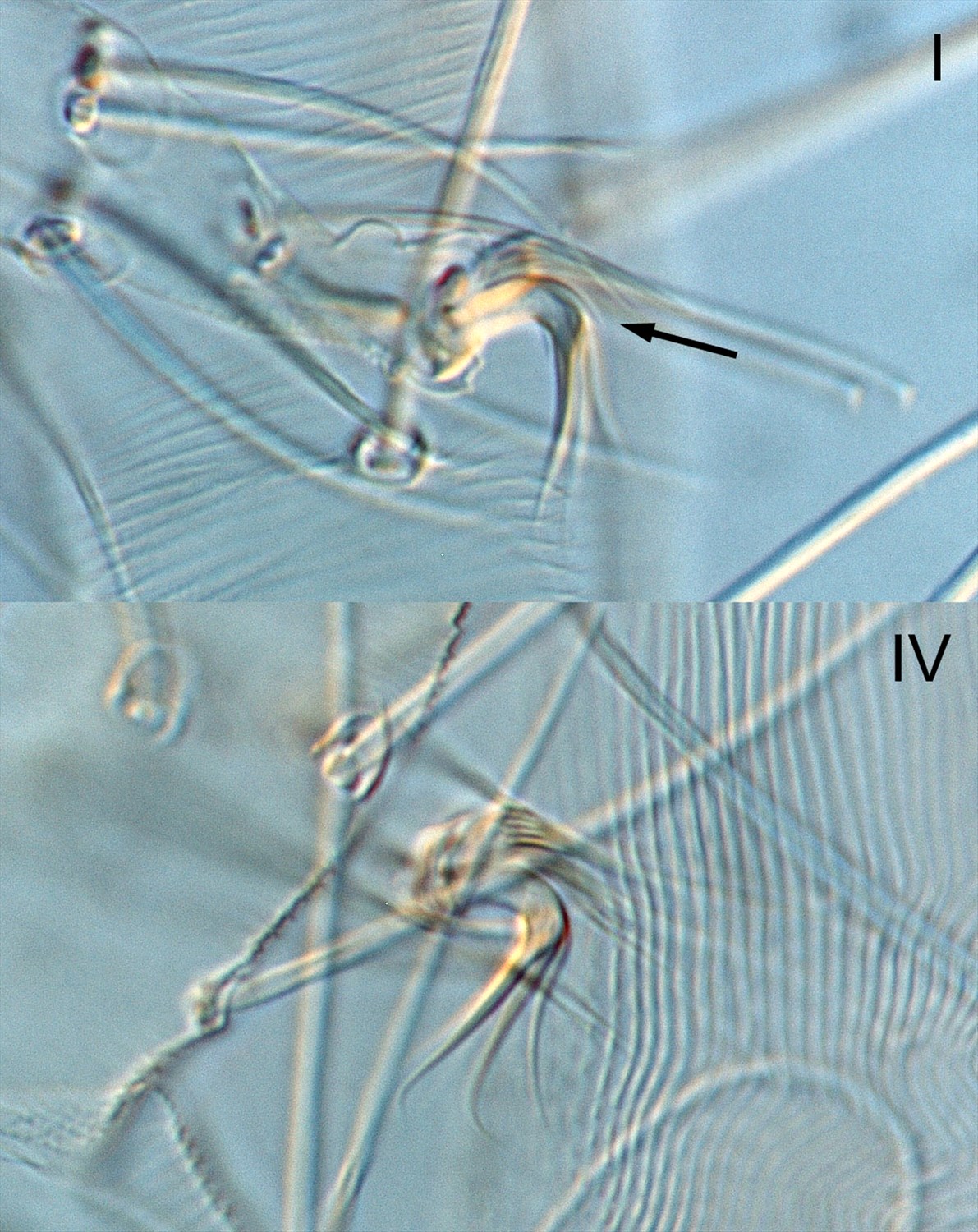
Fig. 1. Tetranychus kanzawai adult female (nontype; Japan) - detail of tarsi I and IV (arrow indicates minute dorsal spur on empodium I).
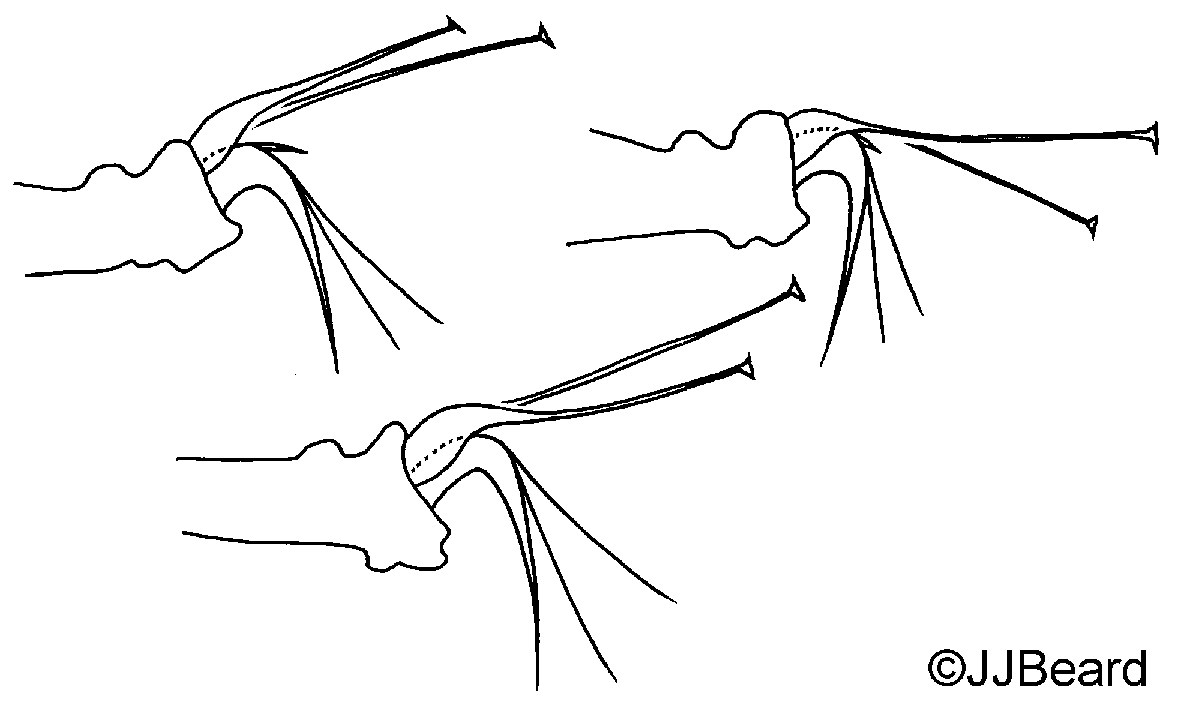
Fig. 2. Tetranychus kanzawai adult female (non-types) - detail of empodia, with minute dorsal spur present or absent.
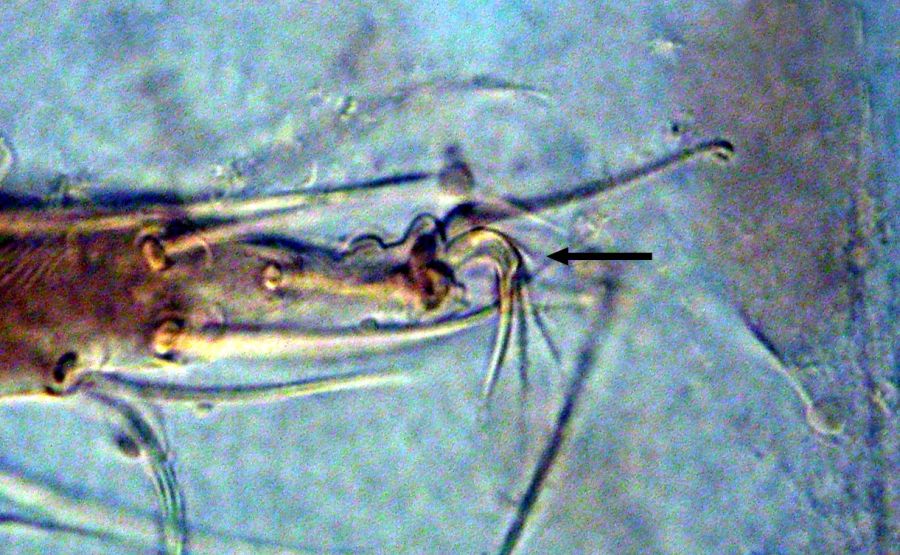
Fig. 3. Tetranychus kanzawai adult female (non-types) - detail of empodia II (arrow indicates minute dorsal spur).
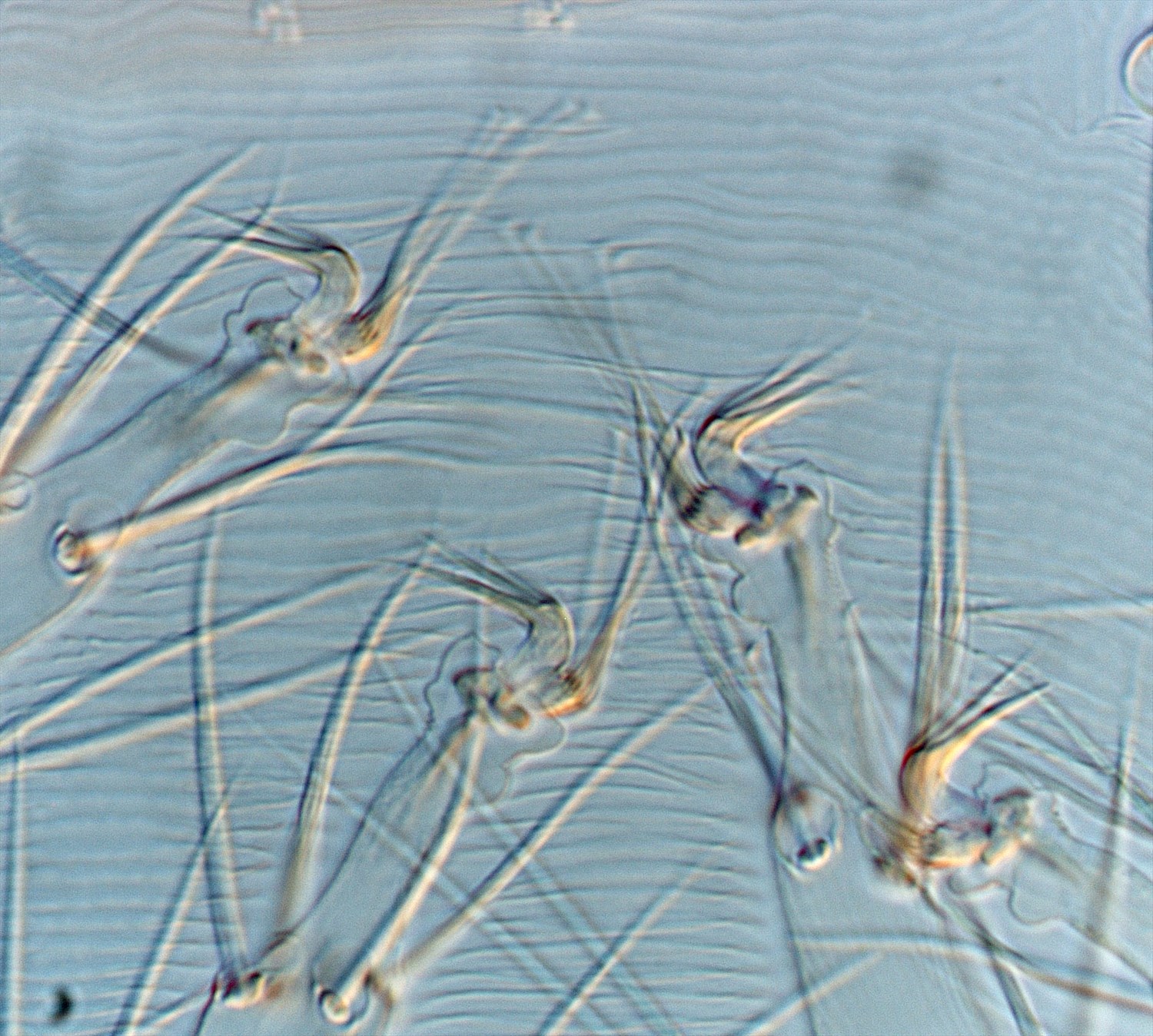
Fig. 4. Tetranychus kanzawai adult female (nontype; Japan) - detail of empodia III and IV (n.b. dorsal spur absent).
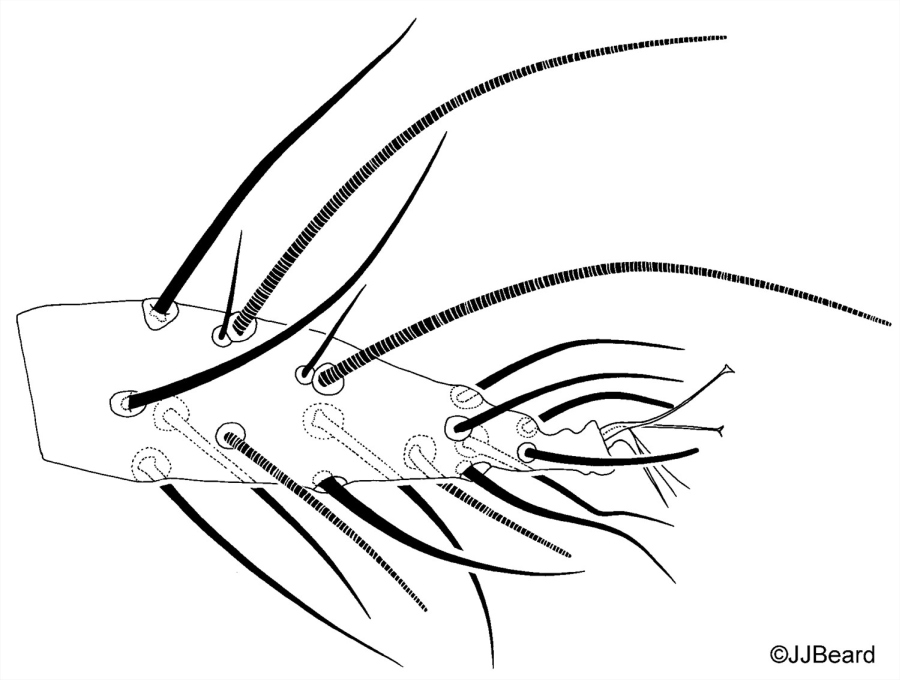
Fig. 5. Tetranychus kanzawai adult female (non-types) - detail of number of proximal setae on tarsus I.
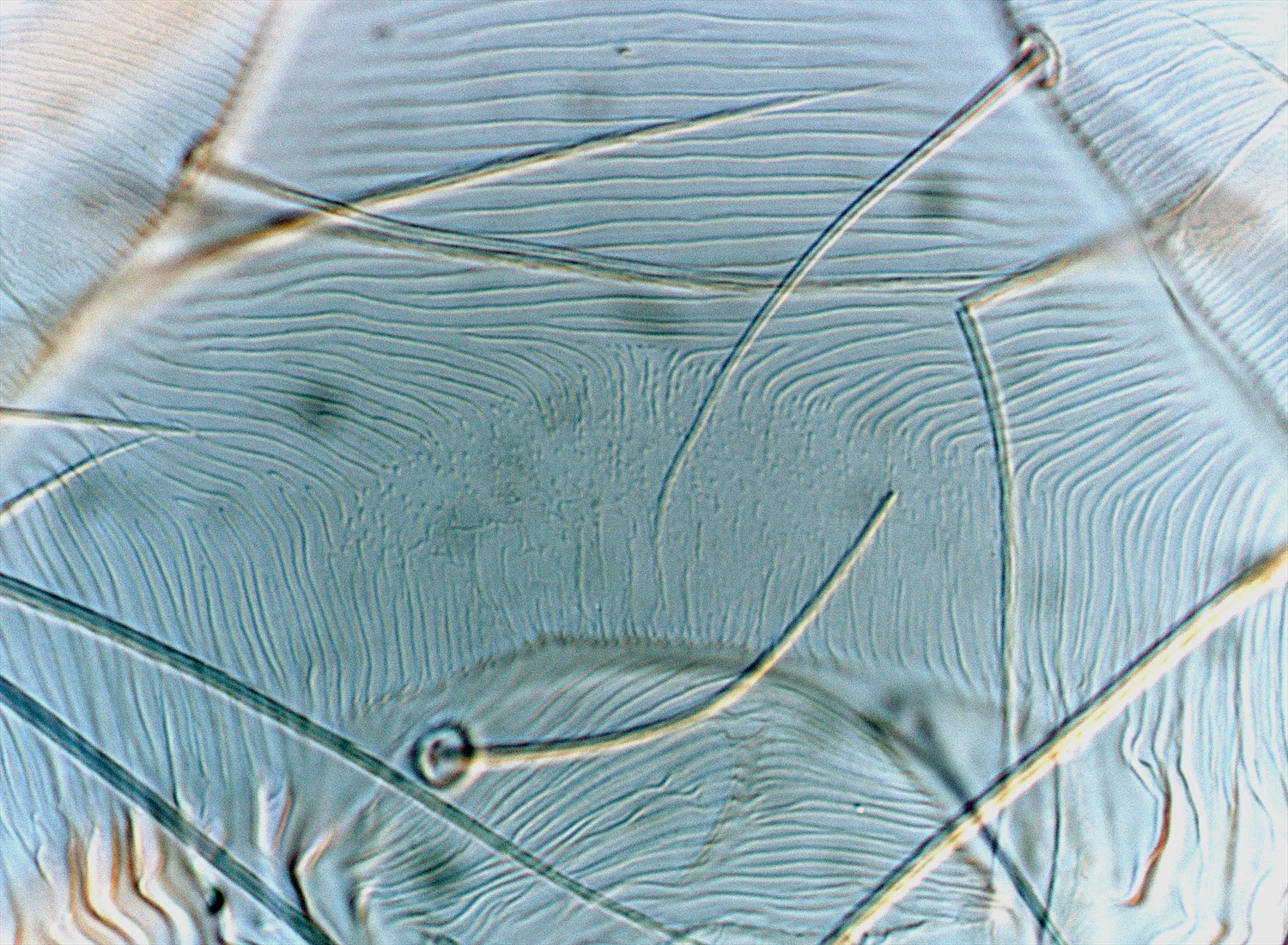
Fig. 6. Tetranychus kanzawai adult female (nontype; Japan) - detail of the pattern of pregenital striae.

Fig. 7. Tetranychus kanzawai adult female (nontype; Japan) - detail of the pattern of pregenital striae.

Fig. 8. Tetranychus kanzawai adult female (nontype; Japan) - detail of peritreme.

Fig. 9. Tetranychus kanzawai adult female (nontype; Japan) - detail of pattern of striae on prodorsum.
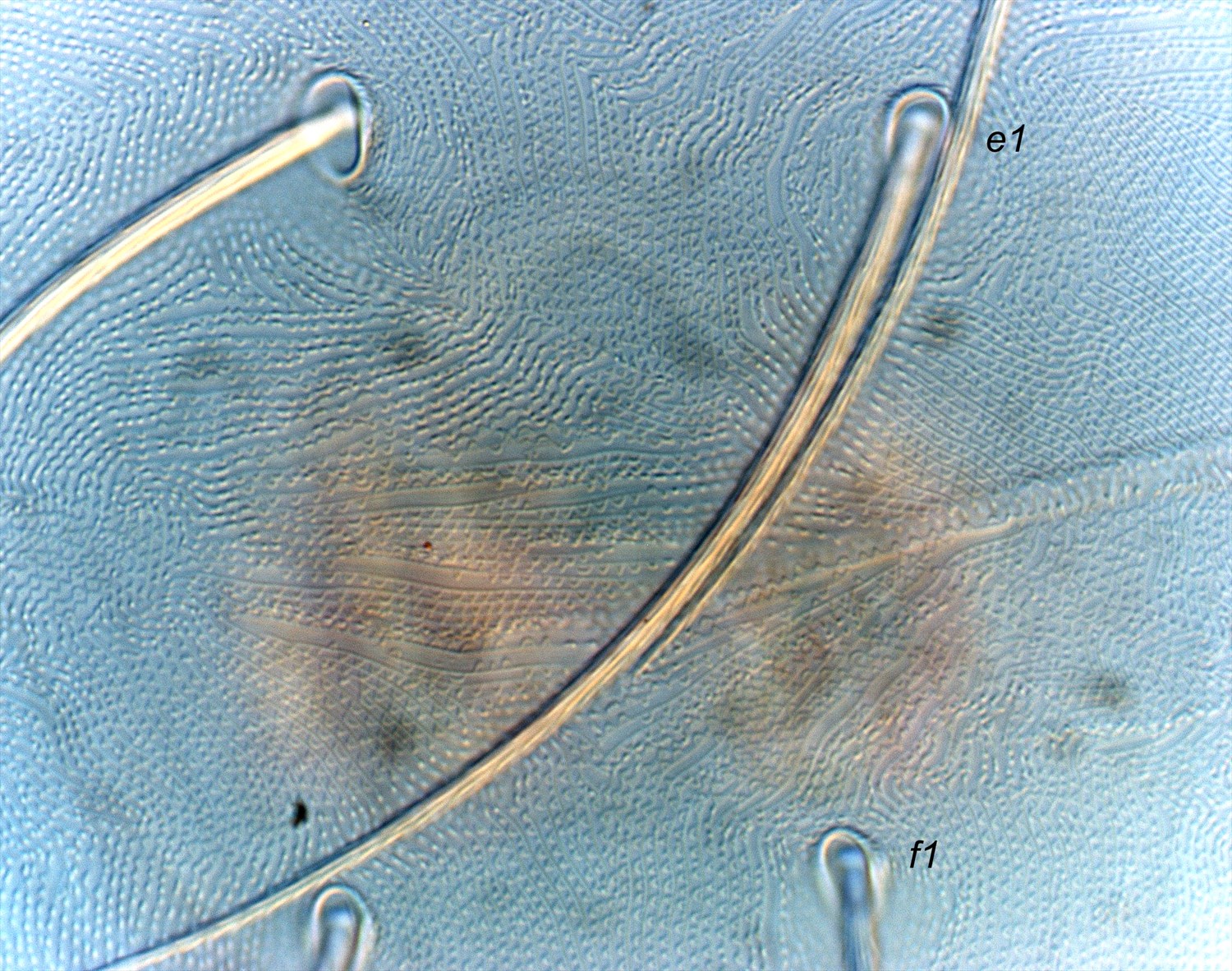
Fig. 10. Tetranychus kanzawai adult female (nontype; Japan) - detail of pattern of dorsal striae between opisthsomal setae e1 and f1.
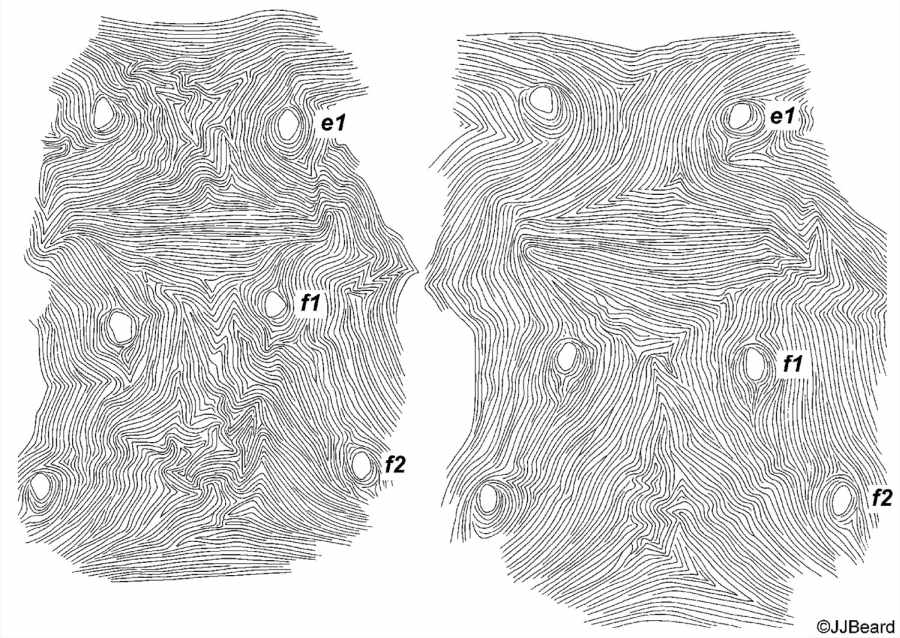
Fig. 11. Tetranychus kanzawai adult female (non-types) - detail of the pattern of dorsal striae between setae e1, f1 and f2.
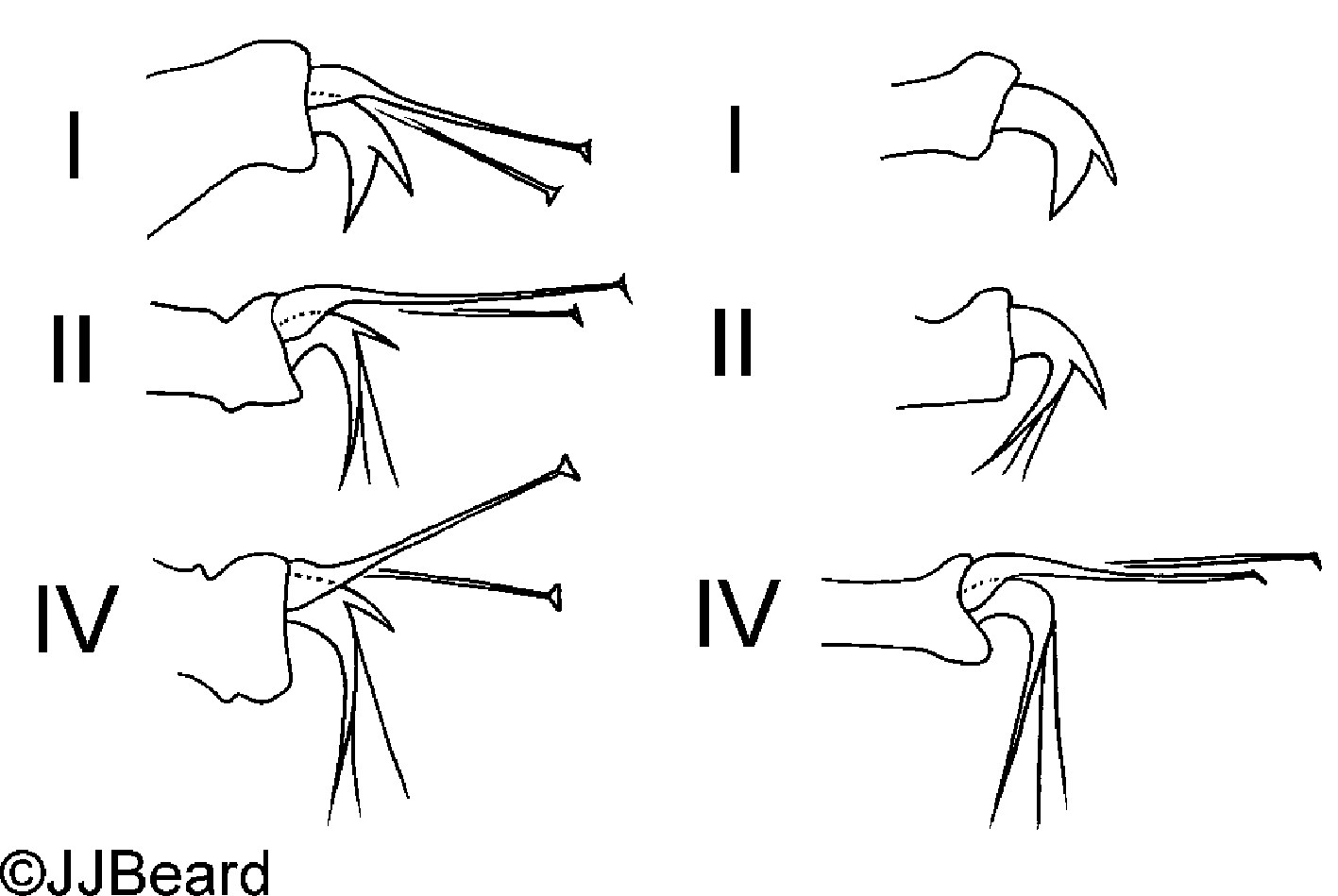
Fig. 12. Tetranychus kanzawai adult male (non-type) - detail of empodia I, II and IV.
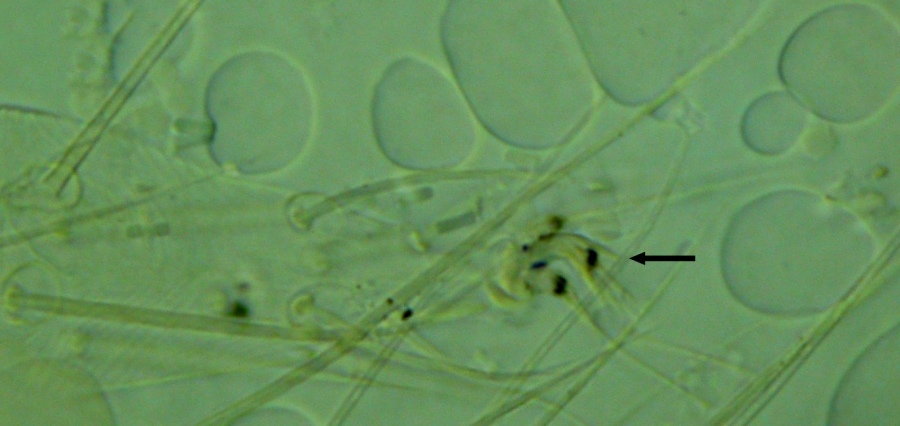
Fig. 13. Tetranychus kanzawai adult male (non-type) - detail of empodium II (arrow indicates minute dorsal spur).
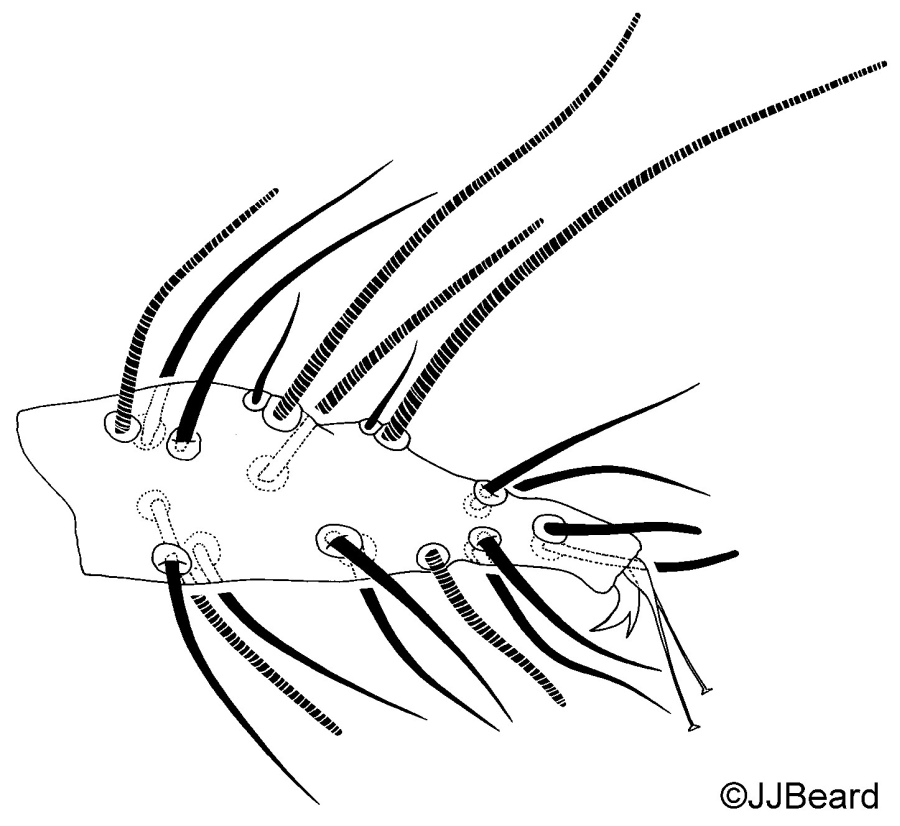
Fig. 14. Tetranychus kanzawai adult male (non-type) - detail of number of proximal setae on tarsus I.
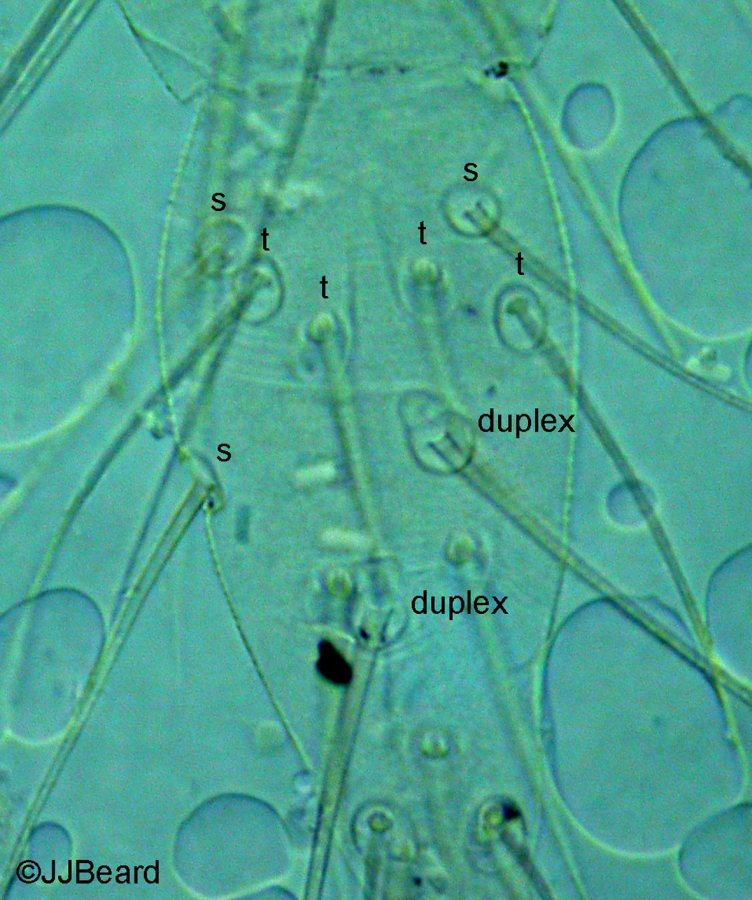
Fig. 15. Tetranychus kanzawai adult male (non-type) - detail of number of proximal setae on tarsus I.
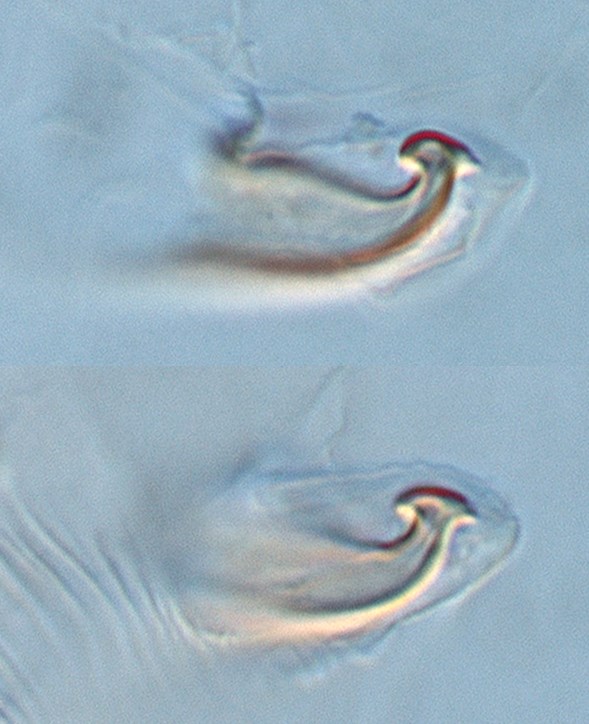
Fig. 16. Tetranychus kanzawai adult male (nontype; Japan) - detail of aedeagus (same individual).
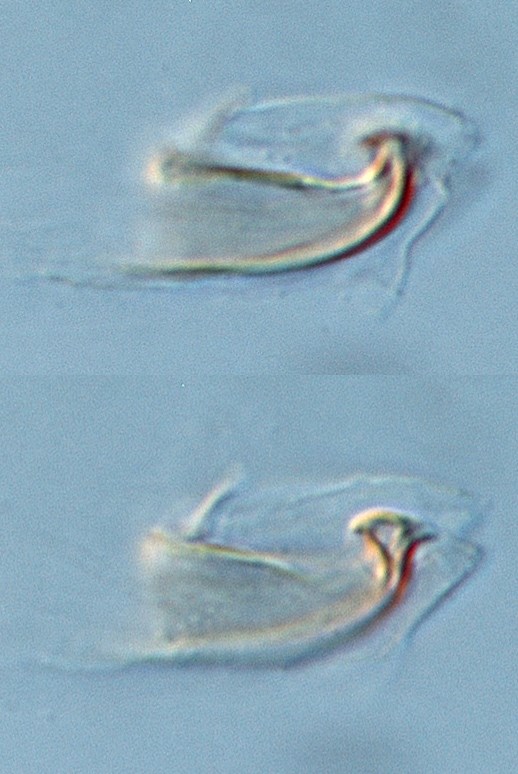
Fig. 17. Tetranychus kanzawai adult male (nontype; Japan) - detail of aedeagus (same individual).
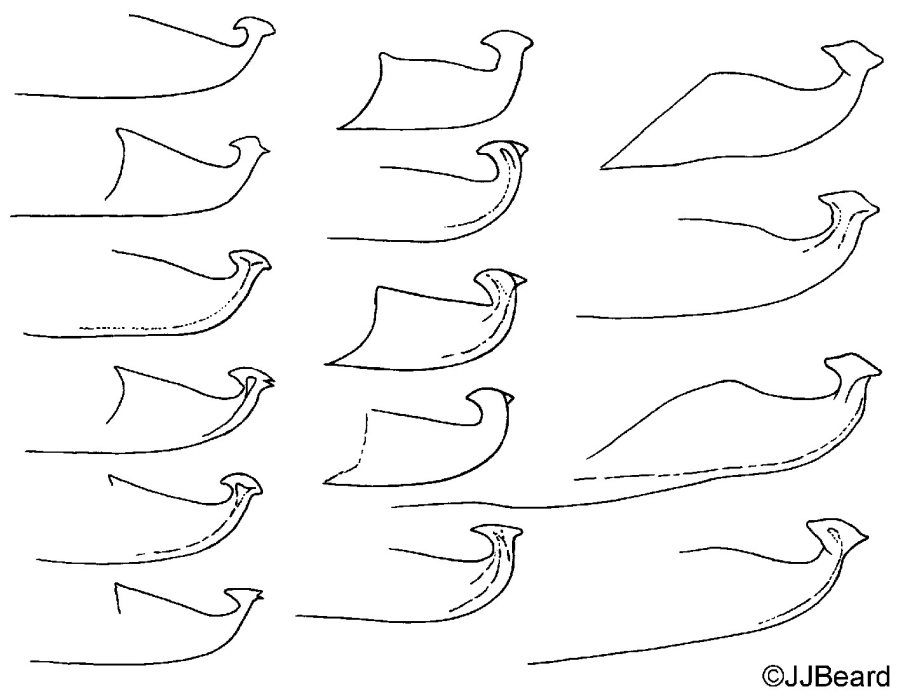
Fig. 18. Tetranychus kanzawai adult males (non-types) - detail of variation in aedeagus between individuals, and within an individual (at different focal points; three different individuals).
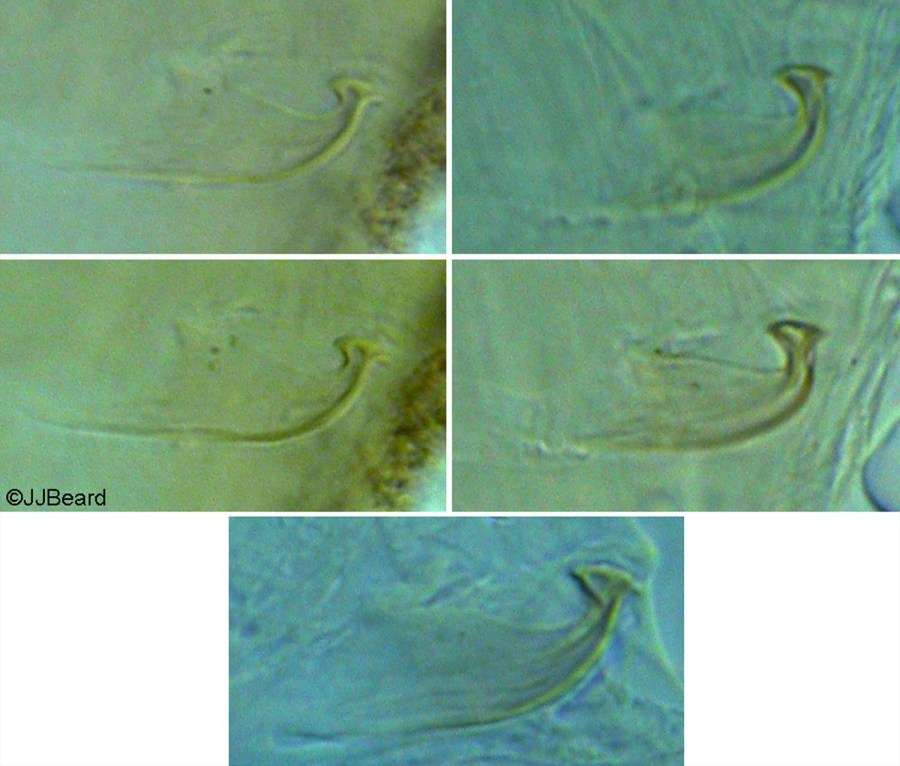
Fig. 19. Tetranychus kanzawai adult males (non-types) - detail of variation in aedeagus between individuals.
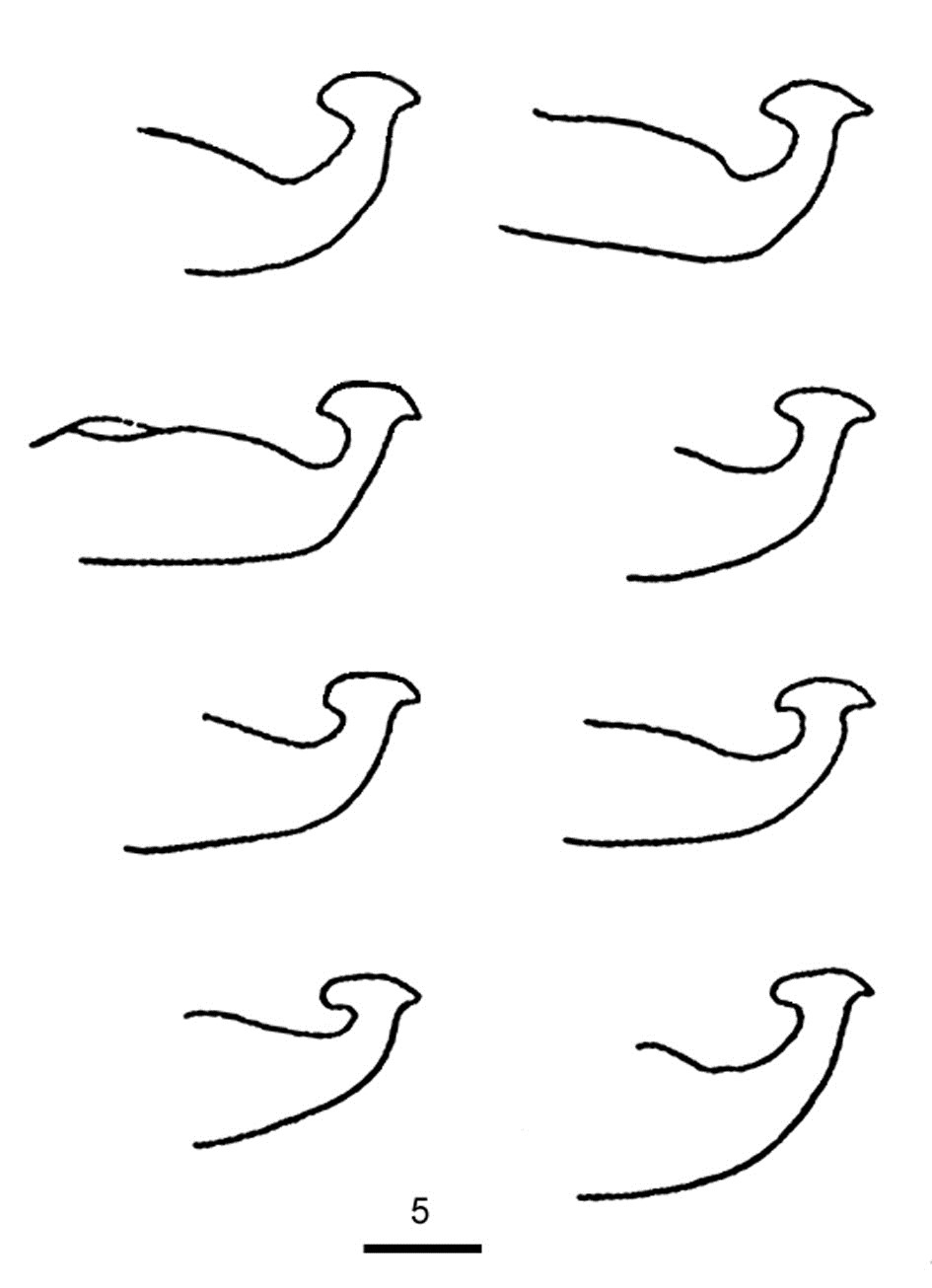
Fig. 20. Tetranychus kanzawai adult male - detail of variation in aedeagus between individuals from Japan (modified from Ehara 1956).
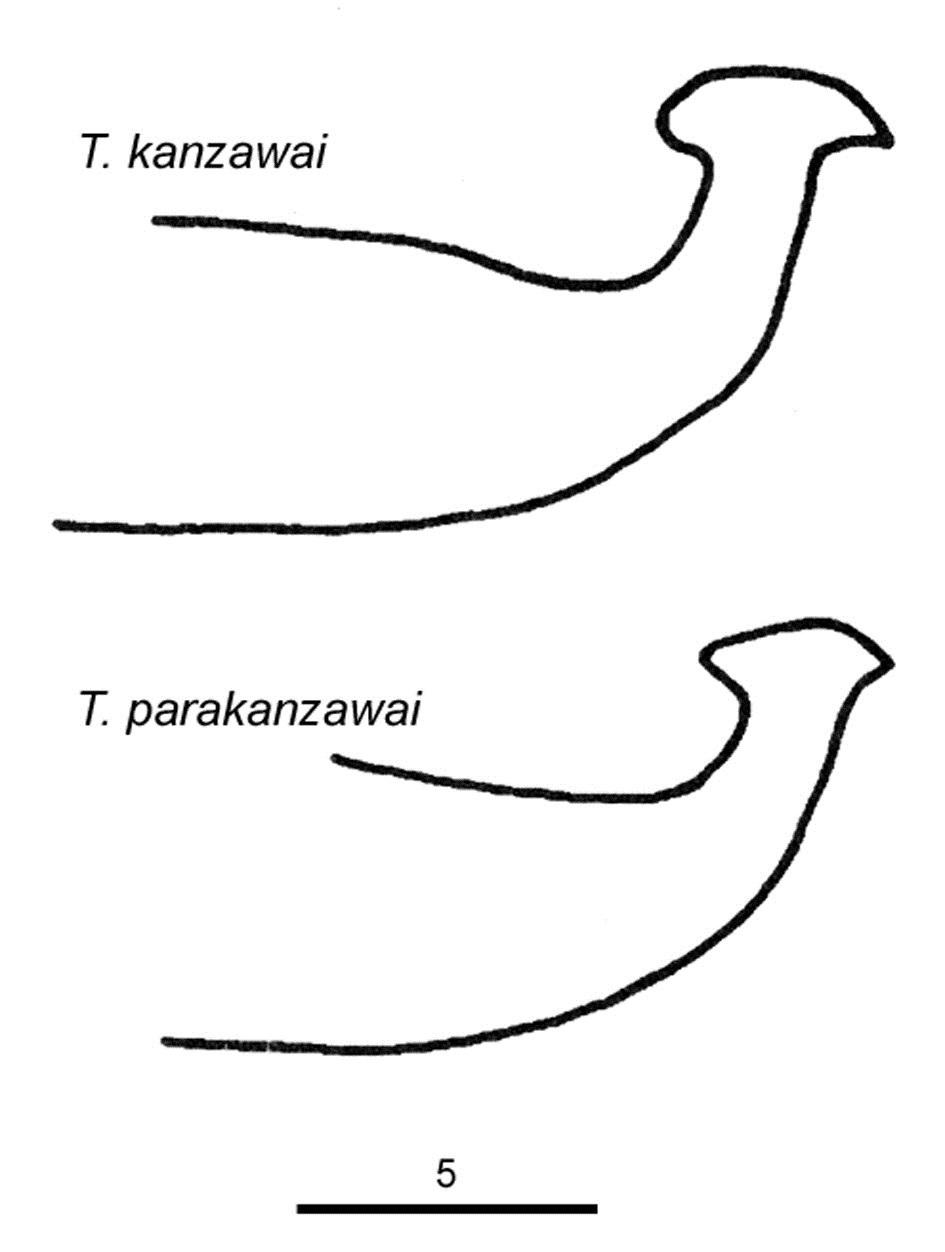
Fig. 21. Comparison of the aedeagi of the two cryptic species Tetranychus kanzawai and T. parakanzawai (modified from Ehara 1999).
Material examined
non-types
Taxonomy
Subfamily Tetranychinae
Tribe Tetranychini
Common Name
- Kanzawa spider mite
- Tea red spider mite
Distribution
+Australia, China, Colombia, Congo, Greece, Hong Kong, India, Indonesia, *Japan, Korea, Malaysia, Mexico, Okinawa Island, Papua New Guinea, Philippines, South Africa, Taiwan, Thailand, USA
Taxonomy Changes
Tetranychus kanzawai Kishida 1927
Tetranychus hydrangeae Pritchard & Baker 1955, synonymy Wainstein 1960
Diagnosis
Female
- empodia I-IV with minute (usually I-II) or absent (usually III-IV) dorsal spur above proximoventral hairs (Figs 1-5)
- tarsus I with the sockets of four tactile setae proximal to, and one solenidion overlapping, the socket of the proximal duplex seta (Fig. 5)
- pregential striae longitudinal, with broad band of broken striae anteriorly; striae can be weak or widely spaced (Figs 6, 7)
- peritreme with long recurved hook (Fig. 8)
- prodorsum with longitudinal striae with strong lobes (Fig. 9)
- dorsal striae between dorsal setae e1-f1 forming the classic diamond pattern - i.e. longitudinal or irregular between e1-e1; transverse between e1-f1; longitudinal between f1-f1 (Figs 10, 11)
- dorsal striae between setae f1-f2 irregular (Fig. 11)
- ventral striae without lobes (Davis (1968) recorded mesal ventral striae with broadly triangular to rounded lobes)
- carmine when alive
Male
- empodia I-IV with strong dorsal spur above proximoventral hairs (4 long) (Figs 12-14)
- empodium I with proximoventral hairs fused to form a ventral claw; dorsal spur much smaler than ventral claw (Figs 12, 14)
- tarsus I with the sockets of four tactile setae and two solenidia proximal to, and one solenidion overlapping, the socket of the proximal duplex seta (Fig. 15)
- shape of aedeagus in individuals identified as T. kanzawai is quite variable, between individuals and within a single individual (as the shape changes with the focus) -
- Japanese specimens: aedeagus dorsally directed, with strong neck and distinct knob; anterior projection broadly rounded (but can appear ventrally truncate); posterior projection short triangular; dorsal margin of knob strongly convex; dorsal margin of shaft at 10-20° angle to ventral margin, abruptly bending dorsad at 70-80° angle; ventral margin of shaft straight (Figs 16, 17, 20)
- other specimens: aedeagus directed dorsally with strong neck and distinct knob; anterior projection short broad triangular; posterior projection short tapering triangular, slightly curved ventrally; dorsal margin of knob strongly convex at one focal point or angulate at different focal point; dorsal margin of shaft at approx. 20° angle to ventral margin before abruptly bending dorsally and then anteriorly to form anterior projection; ventral margin of shaft straight (Figs 18, 19)
- yellow-orange
Hosts
Over 100 recorded species of host plant, including: Ageratum sp., Ambrosia ambrosioides (Asteraceae), Artocarpus heterophyllus (Moraceae), Averrhoa carambola (Oxalidaceae), Boehmeria nivea (Urticaceae), Camellia sinensis (Theaceae), Capsicum annuum (Solanaceae), Carica papaya (Caricaceae), Citrullus lanatus (Cucurbitaceae), Citrus grandis, C. limon (Rutaceae), Cordyline fruticosa, C. terminalis (Asteliaceae), Cucumis melo, Cucurbita sp. (Cucurbitaceae), Euphorbia pulcherrima (Euphorbiaceae), Ficus religiosa (Moraceae), Fragaria sp. (Rosaceae), Gardenia jasminoides (Rubiaceae), Glycine max (Fabaceae), Humulus japonicus, H. lupulus (Cannabaceae), Hydrangea hortensia, H. macrophylla (Hydrangeaceae), Malus domestica (Rosaceae), Manihot esculenta, M. glaziovii, M. maritima (Euphorbiaceae), *Morus alba (Moraceae), Murraya panniculata (Rutaceae), Phaseolus lunatus, P. vulgaris (Fabaceae), Prunus avium, P. campanulata, P. persica (Rosaceae), Pueraria lobata, P. montana (Fabaceae), Pyrus communis, Rosa sp., Rubus crataegifolius, R. thunbergii (Rosaceae), Saccharum officinarum (Poaceae), Solanum nigrum, S. torvum (Solanaceae), Sorghum bicolor (Poaceae), Tectona grandis, Verbena hortensis, V. phlogiflora (Verbenaceae), Vigna angularis, V. catiang (Fabaceae), Zea mays (Poaceae)
Similar Taxa
Tetranychus parakanzawai Ehara 1999
Tetranychus kanzawai can be separated from T. parakanzawai by the length of the knob of the aedeagus (according to Ehara (1999)) (Fig. 21):
parakanzawai = 3.3 μm; kanzawai = 4 μm
see also Similar Taxa for T. urticae
Biology
Little has been published on the biology of this mite in Australia. In an anonymous NSW Department of Agriculture Entomology Branch Insect Pest Bulletin it is recorded that the mite may cause severe injury to hydrangeas during early spring, especially in warm, dry situations. Foliage attacked becomes severly mottled and cup-like blisters form on the young leaves. Flower heads are deformed, reduced in size and show an irregular multi-coloured mottled effect.
The species is well studied in Japan. Although tea and hydrangea are recorded as major hosts, T. kanzawai does better on mulberry (Gotoh & Gomi 2003). At 25°C, development from egg to adult ranges from 9.3-12.2 days for females and 8.6-11.6 days for males. Variation is apparantly caused by seasonal changes in host plant quality. In Japan (moderate temperature region), populations of this mite peak in size during spring (May-June in Japan) and in autumn (September-October in Japan) (Gotoh & Gomi 2000).
Tetranychus kanzawai occasionally causes significant damage to crops, nursery plants and plants in urban landscaping (Jeppson et al. 1975), and is listed as a minor pest of citrus (Gerson 2003).
References
Anonymous (1969) Two-spotted mite and red spider, Tetranychus spp. New South Wales Department of Agriculture Entomology Branch Insect Pest Bulletin 37:1-6
Bolland, H.R., Gutierrez, J. and Flechtmann, C.H.W. (1998) World catalogue of the spider mite family (Acari: Tetranychidae). Brill Academic Publishers, Leiden, 392 pp.
+Davis, J.J. (1968) Studies of Queensland Tetranychidae. III. Records of the genus Tetranychus . Queensland Journal of Agricultural and Animal Science 25:57-67
Ehara, S. (1999) Revision of the spider mite family Tetranychidae of Japan (Acari, Prostigmata). Species Diversity 4: 63-141
Gotoh, T. and Gomi, K. (2000) Population dynamics of Tetranychus kanzawai (Acari: Tetranychidae) on hydrangea. Experimental and Applied Acarology 24 :337-350
Gotoh, T. and Gomi, K. (2003) Life-history traits of the Kanzawa spider mite Tetranychus kanzawai (Acari: Tetranychidae). Applied Entomology and Zoology 38:7-14
Jeppson, L.R., Keifer, H.H. and Baker, E.W. (1975) Mites injurious to Economic Plants. Berkely: University of Californai Press.
*Kishida, K. (1927) Notes on Tetranychus kanzawai n. sp. a new tetranychid mite injurious to leaves of the mulberry tree in Japan. Zool. Mag. 39: 105-107
Migeon, A. and Dorkeld, F. (2006-2017) Spider Mites Web: a comprehensive database for the Tetranychidae. http://www.montpellier.inra.fr/CBGP/spmweb
Notes
Davis (1968:58) recorded this species as Tetranychus hydrangeae Pritchard & Baker; and noted that although the species was recorded only from the Nambour district, the host plant Hydrangea macrophylla is a popular introduced garden plant which has been widely distributed in the country, most likely taking the mite with it.
It should be noted from the images provided for this species that there is morphological variation within this species. Images from three different species of host plant have been presented to demonstrate this variation. It is not known if this variation is significant in terms of species limits or not (i.e. there could be more than one species being called T. kanzawai).
Copyright © 2018. All rights reserved.