General grass terminology
The growth form of most Triodia species is unusual in that the long-lived vegetative parts produce new branches of vegetative growth near the tips, so the new growth just mounds on top of the old, to form a hummock (a ‘small hill’). In a few species new growth is mainly from dense branching only near the base, to form a tuft or tussock. Many species (especially in the Soft group) form a tussock for the first few years of growth and become more hummock-forming in older plants. Here the term hummock is applied to all Triodia species except the few that always maintain a tussock form.


The vegetative parts of a Triodia hummock are composed of culms (the technical term for a grass ‘stem’) that have leaves in the basal part, and (under optimal conditions) terminate in an inflorescence. Irregular (adventitious) branches are formed along the leafy parts of the culm; these may or may not produce roots at their nodes. Some adventitious branches can be almost horizontal, usually produce a tight cluster of unusually short leaves and a dense cluster of roots, and are then called stolons. Most species of Triodia produce stolons, but they are often produced only within or at the margins of the existing hummock, so are inconspicuous. In other species the stolons exceed the existing hummock margin and produce a cluster of surrounding daughter plants. In very old and large hummocks the central part of the plant dies and collapses, and the continuous slow march of living stolons at the margin produces characteristic rings (these may exceed 50 m across in extreme cases).
Triodia leaves have three parts, the blade, sheath and ligule. The sheath is the lower part of the leaf that wraps around the culm (‘stem’). At the apex of the sheath is the needle-like leaf blade proper. The sheath narrows gradually or abruptly at its junction with the divergent blade; the upper side of this junction area is called the orifice. Around the orifice, especially on the upper sheath margins, there are typically distinctive hairs, and this provides one of the most diagnostic features. In some species the base of the leaf blade is contracted into a narrower, paler, tougher stalk called a pseudopetiole. At the junction of the blade and sheath, on the inner surface, is a small transverse band of hairs, sometimes forming a united membrane at the base, called a ligule.

Leaves are divided into a basal sheath and the leafy blade (in some species the base of the blade is modified into a pale pseudopetiole). The point of divergence of the blade from the sheath is called the orifice, which is usually provisioned with hairs.
Triodia leaf blades have modifications for arid living compared to a ‘typical’ flattened grass leaf. The whole blade is hinged along its midline, so that it curls into a tight needle-like structure when dry; in some species the hinge will open to expose the upper surface, while in other species the leaf remains in its needle-like state even in good growing conditions. Stomata (tiny pores in the leaf surface to allow carbon dioxide in and oxygen out) are protected in longitudinal furrows, called stomatal grooves, on both the upper and lower leaf surfaces. The distribution of stomatal grooves on the lower surface is an important character.

The most common inflorescence type in Triodia is a panicle, with lateral branches (which are sometimes further divided. Spikelets diverge from the branches on pedicels.
Grass flowers are arranged into an inflorescence comprising few to many spikelets. Spikelets are dense aggregations of individual tiny flowers sharing a single short axis. Grass flowers are minute and lack nearly all traces of petals and sepals. Flowers are hidden within a series of highly modified leaves (bracts) that have specific names depending on their position. At the base of the whole spikelet is a pair of papery glumes that initially enclose all the flowers – the glumes remain attached to the plant after seed matures and are frequently the only remaining spikelet parts available for identification. In Triodia, each spikelet contains two to many (usually >3) flowers, each one surrounded by two papery bracts known as the lemma and palea. Together the whole unit of lemma+palea+flower is known as a floret. In Triodia, the floret is the normal unit of dispersal, since the seed (technically a caryopsis) remains inside the floret when mature. The florets in a spikelet are arranged in two alternating rows, each successive floret a little smaller than the ones below; the florets at the apex often contain incompletely developed flowers.
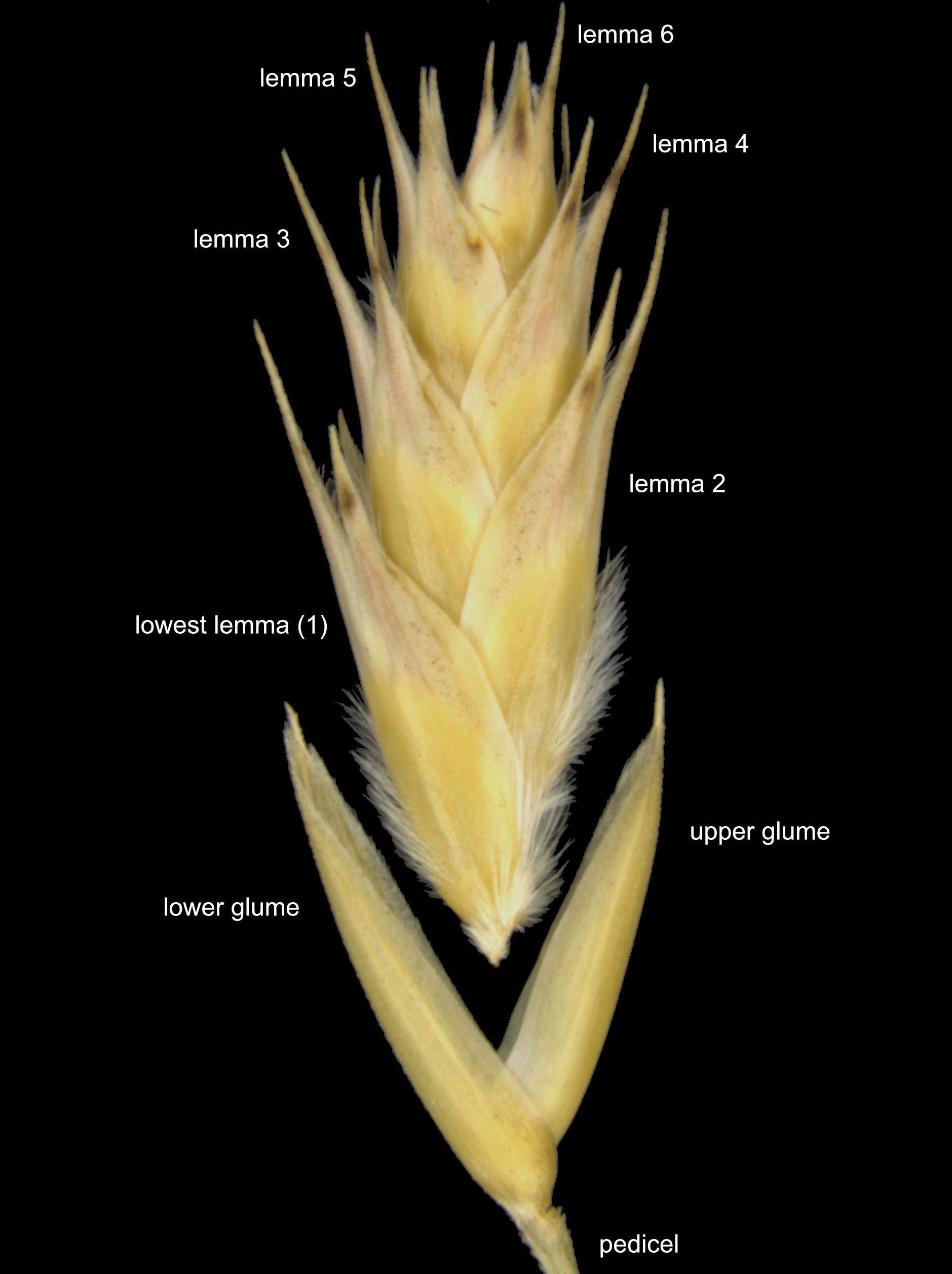
Spikelets are dense aggregations of flowers covered by bracts. A pair of glumes subtend the alternating florets. Flowers and paleas are usually hidden within the lemmas, so at this scale florets and lemmas appear interchangeable. Florets and lemmas are numbered from the base. The basal-most lemma (and palea) is the one usually measured.
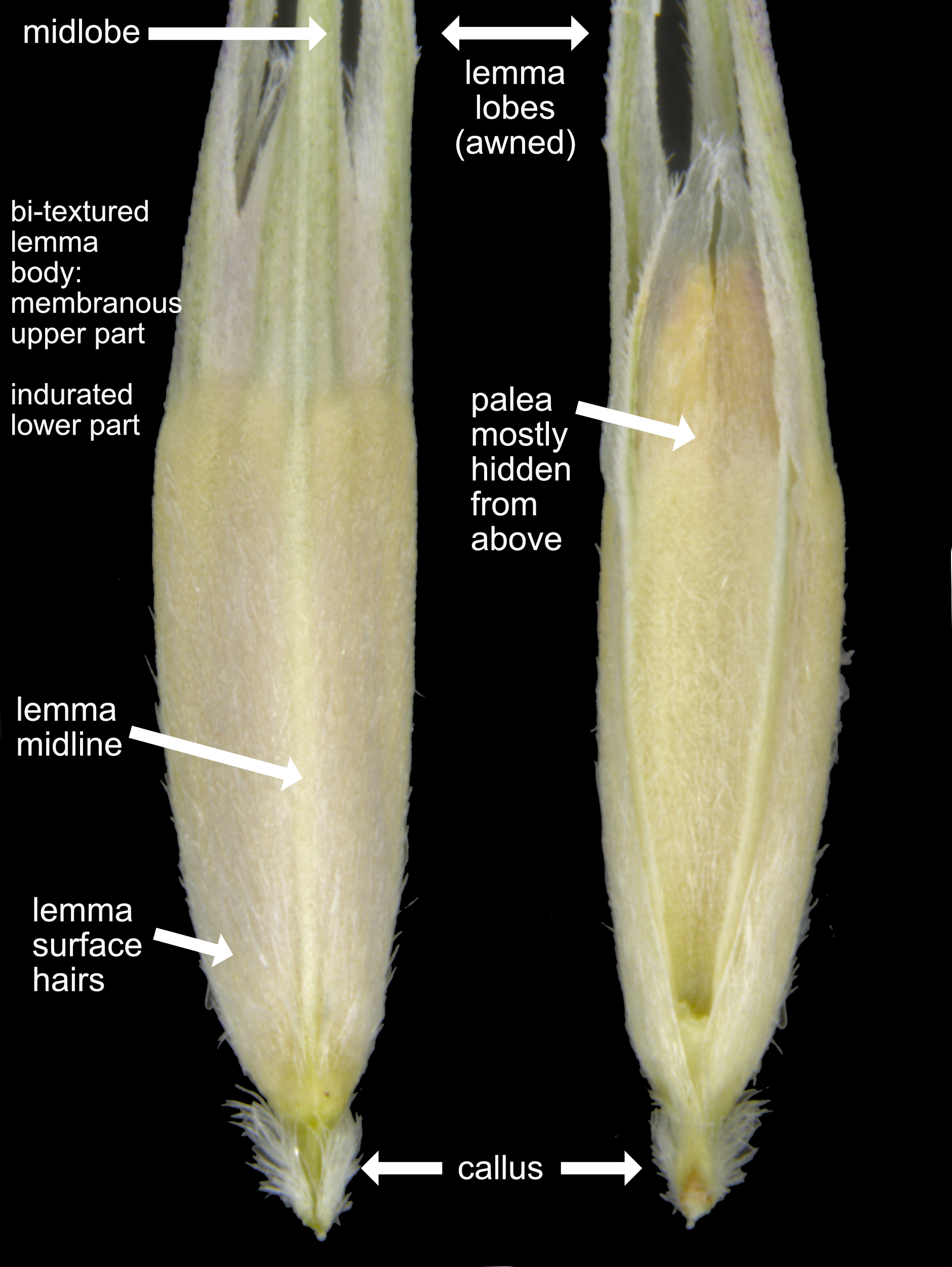
Each floret is composed of a large lemma and a smaller palea hidden underneath the lemma, and usually only visible with dissection of the spikelet. The flowers are hidden between the lemma and palea. The lemma has a thickened callus at the base, and lobes at the apex that are sometimes drawn out into long, thread-like awns. Important characters on the lemma body are the nature and distribution of hairs relative to the midline, and whether the lemma texture is uniform or has a sharp texture transition (bitextured).
Inside each (fertile) floret are the tiny floral parts (these are not particularly useful for identification purposes). The flower itself comprises three anthers containing the pollen, an ovary topped by two short feathery styles, and two minute scales known as lodicules. After fertilisation, the ovary swells and develops into a small fruit (grain) called a caryopsis that has a very thin dry wall and contains a single seed. Since the fruit wall never splits, and the caryopsis is structurally nearly all composed of the seed, the term seed is in practice interchangeable with caryopsis, and is used by most practitioners (although technically incorrect).

If lemmas or paleas are removed, the minute flower inside is exposed, revealing the familiar flower organs of a single ovary, 2 styles and 3 anthers. The pair of lodicules at the base possibly represent the highly reduced petals or sepals.
Additional features
Palatability to stock is reported on species profiles where known, based on published literature and personal observations (abbreviations MB and BA refer to M. Barrett & B. Anderson respectively). As a general rule, “soft spinifexes” (see Phylogeny below) are palatable as young plants or regrowth in active growth, but become unpalatable as plants toughen up with drought stress and age. In contrast the “hard spinifexes” are unpalatable due to their very tough leaves and sharply pungent leaf tips.
Regeneration strategy
Where known, the observed regeneration strategy is reported, based mainly on Rice & Westoby (1999), in cases where the reported species can be updated to modern taxonomy, and personal observations (abbreviations MB and BA refer to M. Barrett & B. Anderson respectively).
Whether plants resprout following fire (resprouters), or only propagate from seed (seeders) is probably dependent on several factors. Some species appear to completely lack sufficient below-ground storage or regenerating buds to respout; survival of a few individuals that are incompletely burnt is discounted as resprouting behaviour. Other species show abundant resprouting behavior, but often exhibit patchiness in their response. This patchiness may be due to trait variation within species, fire intensity, timing of fire, plant condition prior to burning, or environmental conditions following fire. Post-fire recruitment from seed post-fire is likewise patchy, due to a similar range of variables affecting seed quality and abundance, seed dormancy, germination requirements, emergence and seedling survival.
Phenology
Phenology (timing of reproduction stages) is not reported under species. All Triodia species in the Pilbara start producing inflorescences following substantial wet-season rains in about December or January, flower (release pollen and expose their stigmas) with available rains in January or February, and mature their seed in late February or March. These stages can vary by at least a month each year depending on rainfall. In low-rainfall years populations often produce little or no seed.
When winter rainfall occurs (May-July)
Most Pilbara Triodia species do not usually flower in response to winter (May-July) rainfall, but sometimes just a few plants in a population may produce inflorescences. An exception to this are a few species in the Basedowii group, such as T. basedowii and T. vanleeuwenii, which will sometimes produce inflorescences following exceptional winter rains. However, the full range of species that are able to respond to winter rainfall is not known.