
T. wiseana spikelet of short-lobed form.

T. wiseana habit on low rise over rock / gravel.

T. wiseana orifice of typical form.

T. wiseana leaves and orifice of exceptionally hairy form.

T. wiseana orifice of minimally hairy form.
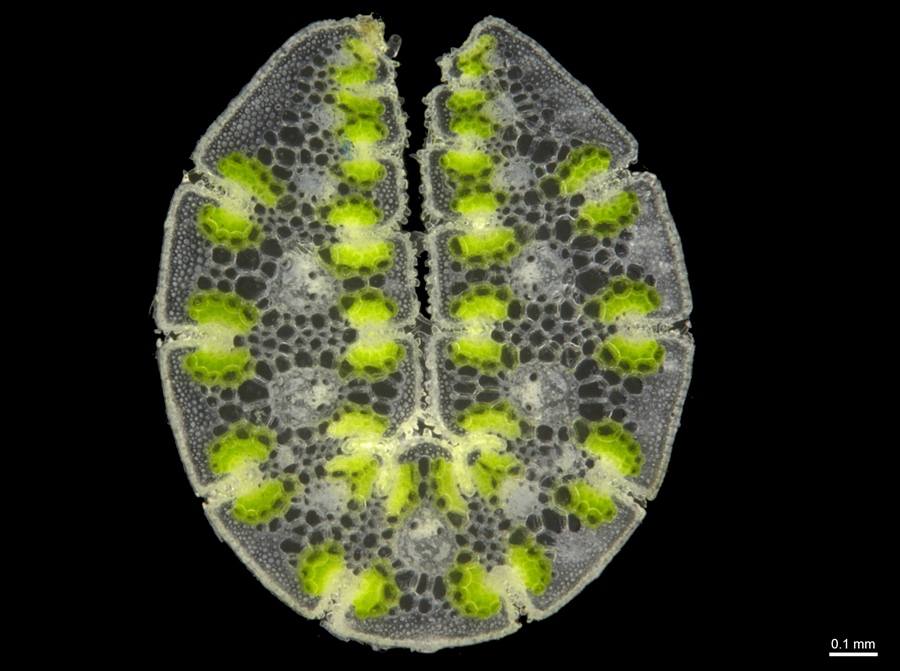
T. wiseana leaf section.

T. wiseana spikelet of long-lobed form.

T. wiseana glumes.

T. wiseana lemmas of short-lobed form.

T. wiseana lemma of long-lobed form.

T. wiseana paleas.
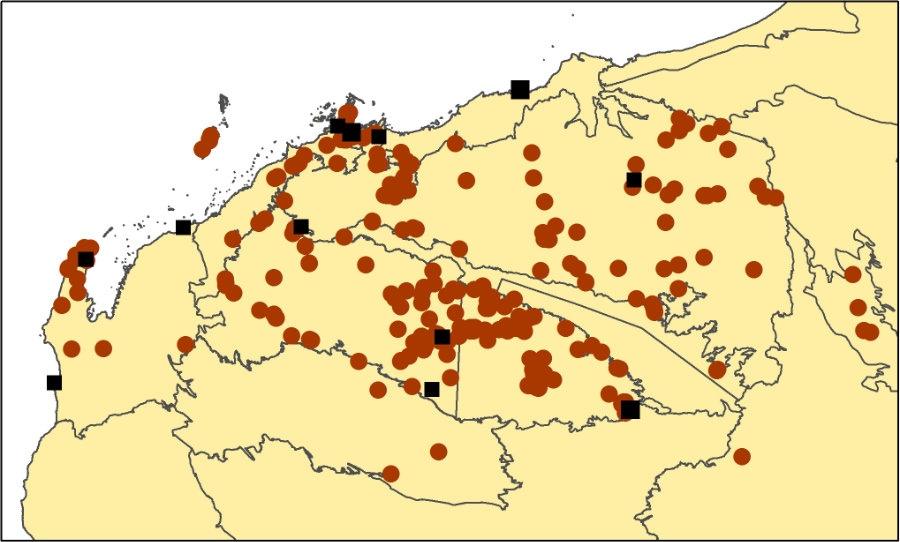
T. wiseana map.
Name
Triodia wiseana C.A. Gardner
Citation
J. Roy. Soc. Western Australia 27: 166 (1942)
Derivation
wiseana — in honour of Frank J. S. Wise (1897–1986), Australian agricultural scientist and later Premier of Western Australia.Common name
Stiff-haired Spinifex, Limestone Spinifex
Synonyms
None
Diagnostic features
Foliage non-resinous; leaf sheath surfaces hairy or not; leaf blades amphistomatous (hard-type); orifice hairs stiff, straight or bent in age, 2–7 mm long, nearly always extending onto at least the base of the leaf blade; inflorescences branched; longest basal pedicels 2–11 mm long; lower glume 3–5-nerved; lowest lemma lobed, midlobe 0.6–4.1 mm long, not awned; lemma body with hairs along midline and lower margin; occurs on rocks or gravel, or very rarely on flat loam apparently lacking rock, throughout the Pilbara bioregion, Cape Range, and very isolated localities in adjacent regions.
Habitat
Occurs on rocky hills and slopes, or gravel rises and flats, and very rarely on seemingly open plains (e.g. near Pyramid Hill SE of Karratha).
Distribution and frequency
Endemic to north-western Australia. Occurs throughout the Pilbara region, also Cape Range and a few scattered locations in the Murchison and Great Sandy Desert bioregions. Usually on rocky and gravelly substrates, including banded iron, limestone, dolomite and shale. Absent from nearly all alluvial, sand and loam plains, but a large population occurs on the plains around Pyramid Hill east of Karratha. Only occurs on rocky/gravelly margins of the Fortescue Valley.
Similar species
Triodia wiseana belongs to the Angusta-Wiseana group, by virtue of having amphistomatous (hard-type) leaf blades and 1–3-nerved glumes.
No other Pilbara species has the characteristic stiff hairs extending on to the leaf blade (although some species such as T. brizoides and T. spicata have woolly hairs extending onto the blade, and a few species have stiff hairs restricted to the orifice). However, in some populations of T. wiseana, the ±straight hairs are restricted to the orifice, and then can be similar to T. angusta, T. longiceps or T. longiceps X wiseana hybrids.
Triodia wiseana is normally easily distinguished from T. angusta by the stiff hairs that are usually present on leaf blades of T. wiseana, but rare forms with hairs restricted to the orifice are very similar to some forms of T. angusta. Triodia wiseana lacking hairs on the blade can be distinguished from T. angusta by having longer basal pedicels >5 mm long and irregular spikelet arrangement (pedicels <3 mm long and spikelet with a secund arrangement on panicle branches in T. angusta).
Triodia longiceps lacks hairs, or have only minute hairs at the orifice (T. wiseana always has some long hairs at least at the orifice).
Triodia longiceps X wiseana hybrids can be distinguished from most forms of T. wiseana (including known co-occurring forms) by having hairs restricted to the orifice (longer stiff hairs extending onto at least the base of the leaf blade in most T. wiseana). Rare forms of T. wiseana with hairs restricted to the orifice always have prominent lemma lobes 1/3–1/2 of the total lemma length (T. longiceps X wiseana lemma lobes are minutely notched for less than 1 mm).
Conservation status
Not considered at risk.
Identification without florets
The notes under Similar species above cover material without florets.
Variation
Triodia wiseana is a very variable species. The indumentum, although always of ± straight hairs, varies from a few hairs at the apex of the sheath margins (the orifice), to spread over the entire sheath and blade.
Some features, although usually fairly uniform within populations, can differ substantially between different parts of the distribution. In particular, the lemma lobes can be short or long, with a strong geographical correlation (See below). Plants with few hairs near the orifice are only found in the west Pilbara, e.g. near Mt Stuart Station, and south and east of Marble Bar.
Plants with both few hairs AND long lemma lobes, i.e. those from the far west Pilbara and south-east of Marble Bar, were placed under variety breviloba by Burbidge (1946a, 1953), contrasting with the type form from Mt Margaret Pass (near Mt Sheila Station), which belongs to the Hamersley Range, short-lemma-lobe form. However, the features of hairs and lemma lobing is more variable than was known to Burbidge, since many populations have abundant leaf hairs and long lemma lobes.
Genetic markers (Genotype-by-Sequencing, Barrett et al. in prep.) show four genetic groups across the Pilbara, that intergrade in the central-western Pilbara: (1) Cape Range, (2) North and East Pilbara, from Millstream east, (3) west Pilbara, west of about Millstream and Rocklea Station, and (4) Hamersley Range from Rocklea and Mt Sheila Stations east to Newman.
Group (4) throughout most of the Hamersley Range has short lemma lobes (midlobe of lowest lemma 0.6–1.3 mm long), while groups (2) and (3) in the west and north Pilbara mostly have deeply lobed lemmas (midlobe of lowest lemma 1.6–4.1 mm long). Group (1) from Cape Range are intermediate, with midlobe of lowest lemma 1.1–1.8 mm long.
It is possible that a sub-specific classification could be established, in part based on division of the lemma lobes, but since there are a few exceptions to the lemma lobe character, and there is genetic intergradation in the central-west Pilbara, we have not attempted to formalize one.
Polyploid races occur within most of the genetic lineages, and across the different morphological forms.
Notes
The concept of T. wiseana used here (for Pilbara forms) is the same as that of Lazarides (1997), Lazarides et al. (2005) and Ausgrass (Sharp & Simon, 2002; Simon & Alonso, 2014).
A full description of T. wiseana, in the sense used here as endemic to the Pilbara and surrounds, can be found in Burbidge (1953), but see also the notes on variation above.
Plants previously referred to T. wiseana in the Kimberley and Northern Territory are genetically divergent from Pilbara T. wiseana (including the type). Kimberley and Northern Territory ‘T. wiseana’ are genetically closer to T. intermedia, and we consider them to belong to one or more undescribed species. Measurements of T. wiseana used here were made only from plants occurring within the Pilbara and surrounds.
Very rare hybrids with T. wiseana are known (see Triodia longiceps X wiseana species profile).
Triodia wiseana is not palatable to stock.
Different forms have been observed resprouting profusely or to be entirely fire-killed (Rice & Westoby 1999, MB & BA observations). It is not known whether regenerating ability is genetically controlled, environmentally driven (e.g. fire intensity) or both.