
T. epactia spikelet.

T. epactia habitat.
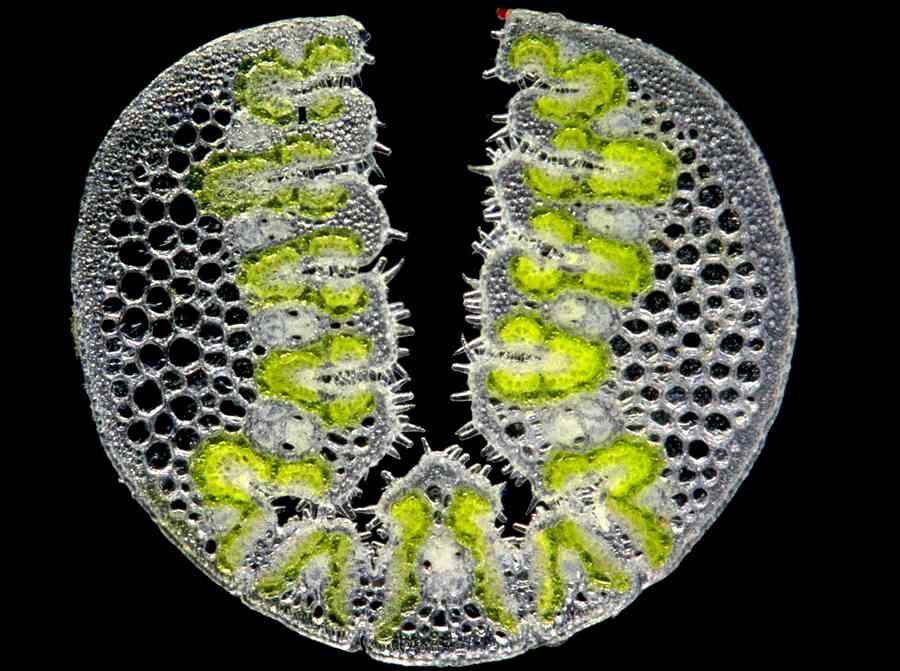
T. epactia leaf section.

T. epactia inflorescence.

T. epactia glumes.

T. epactia lemmas.

T. epactia paleas.
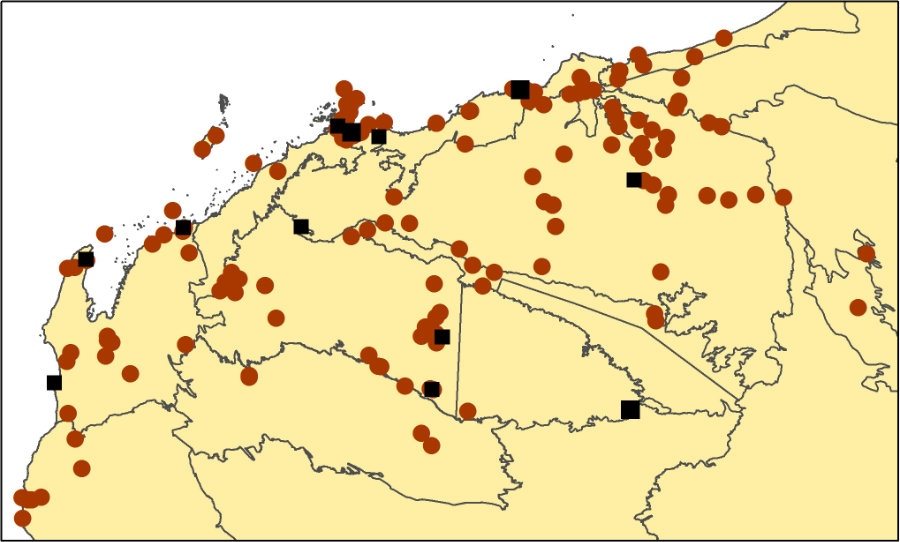
T. epactia map.

T. epactia (right) and T. pungens (left) spikelet comparison; note difference in lobe length and angle.
Name
Triodia epactia S.W.L.Jacobs
Citation
Nuytsia 8: 220–221 (1992)
Derivation
epactia — the Latinised form of the Greek adjective epaktios, coastal, in reference to a maritime habitat (named when it was only known from subcoastal areas).Common name
Humpback Spinifex
Synonyms
None
Diagnostic features
Foliage copiously resinous; leaf sheaths glabrous on surfaces and margins; leaf blades epistomatous (soft-type); spikelets with 3–17 florets; lower glume (3–)4–11 mm long, ovate to broadly lanceolate, L:W 1.8–3.8; lemma lobes (1.2–)1.5–5 mm long, not awned; lemma body strongly indurated and differentiated from membranous lobes at upper margins, often with a slight ‘hump’ in profile at the change in texture; palea indurated, glabrous, winged; distribution across most of north-west Australia, but absent from the eastern Hamersley range south of the Fortescue River and east of Wittenoom.
Habitat
Occurs on most substrates, but particularly extensive on sandy plains and dunes, including coastal sands. Also occurs on loam and clay flats, and rocky hills of ironstone, chert and limestone.
Distribution and frequency
Widespread in north-western Australia. Common throughout most of the Pilbara and surrounds, however it is absent from the eastern Hamersley Range and Little Sandy Desert, where it is replaced by T. pungens. In the south-east Pilbara, T. epactia only occurs west of Wittenoom and Karijini National Park, and east of Wittenoom it only occurs on the north side of the Fortescue River bed.
Similar species
Triodia epactia belongs to the Soft group, sharing epistomatous (soft-type) leaf blades. Several other species with epistomatous (soft-type) leaves and resinous foliage occur in the Pilbara.
Triodia avenoides, T. basitricha, T. melvillei, T. schinzii can be distinguished by having narrowly lanceolate glumes (L:W>5) (ovate to oblong or broadly lanceolate with L:W<4 in T. epactia), and prominently awned lemmas.
Triodia veniciae has hairy leaf sheath surfaces and narrowly lanceolate glumes (L:W 5–7). Only Triodia biflora and T. pungens share epistomatous (soft-type) leaves, resinous foliage and ovate to broadly lanceolate glumes.
Triodia biflora has only 2 fertile florets per spikelet, (and a third minute, vestigial one on a long stalk), and glabrous lemmas that lack lobes or have only minute lobes (T. epactia has 4-many florets per spikelet, usually hairy lemmas and distinct lemma lobes >2 mm long).
Triodia pungens is extremely similar to T. epactia, and the two taxa may be better treated at subspecies rank — see under T. pungens for discussion on the taxonomic difficulties of applying this name to Pilbara forms. Other than by their non-overlapping but complementary distribution (see below under Identification without florets), the two species can only be distinguished by the following features:
T. epactia: Lemma lobes (1.2–)1.5–5 mm long, from anthesis (flower opening) the lobes distinctly ascending-spreading, divergent from each other and the spikelet outline; margin of lateral lemma lobes prominently winged laterally; body of lemma conspicuously indurated, thicker and more yellow than ± membranous lobes, ± bitextured, fully mature lemmas in profile having a distinct ‘hump’ in outline due to the abrupt change in lemma thickness.
T. pungens: Lemma lobes short, 0.6–1.5 mm long, appressed to the next lemma or almost so, and not or scarcely diverging from each other even in age; margin of lateral lemma lobes not winged or prominently winged laterally; body of lemma indurated, but grading more gradually into membranous margins, not or only weakly bitextured (or sometimes more strongly bitextured), in profile the outline smoothly curved and lacking a distinct ‘hump’.
Conservation status
Not considered at risk.
Identification without florets
Triodia epactia, T. biflora and T. pungens are only three Pilbara species that have both resinous foliage and ovate to broadly lanceolate glumes (L:W4) (most other species with resinous foliage have narrowly lanceolate glumes and L:W≥5).
Triodia biflora has glumes 2.5–3.5 mm long [(3–)4–11 mm long in T. epactia], larger inflorescences with more widely spaced spikelets, is only found in gorges, below cliffs or ridges on hills, and is extremely rare within the distribution of T. epactia (see below).
Without florets, T. epactia can only be distinguished from T. pungens using distribution; the two species have complementary distributions, but never approach more than about 10 km from each other. Although T. epactia occurs throughout most of the Pilbara and surrounds, it is absent from the eastern Hamersley Range and Little Sandy Desert. Triodia epactia is replaced by T. pungens from Karijini National Park and Wittenoom east, and south of the Fortescue River bed along the Fortescue Valley.Variation
Triodia epactia is very variable in the number of florets per spikelet, in the length of the lemma lobes, the width of the lateral lobe wing, and glume shape. Burbidge (1946a) noted eight forms in the north Pilbara, based on floret number, hummock size, resin production, stolon production, substrate and palatability to stock. However there appears to be no clear distinction between these variants, and they are often variable within a single population. In rare extreme cases the lemma lobes appear shortly awned.
Notes
The species concept used here is the same as Lazarides (1997), Lazarides et al. (2005) and Ausgrass (Sharp & Simon, 2002; Simon & Alonso, 2014), and the descriptions there are accurate, however maps presented therein are inaccurate since they incorporated many sterile specimens of T. pungens misclassified under T. epactia.
The species limits and non-Pilbara distribution of Triodia epactia remain uncertain. Triodia pungens together with T. epactia appears to be a polyphyletic species complex (see multiple positions of T. epactia and T. pungens in trees in Toon et al. 2015). The Pilbara form of ‘T. pungens’ appears to be much closer to T. epactia than true T. pungens of arid north Queensland and the Northern Territory. Further genetic and morphological studies are needed.
Plants show variable resprouting behavior, sometimes resprouting and sometimes not (Rice & Westoby 1999; MB obs.), possibly due to fire intensity and subsequent rainfall.