
T. veniciae spikelet (shorter extreme).

T. veniciae habitat.

T. veniciae sheaths and orifice.
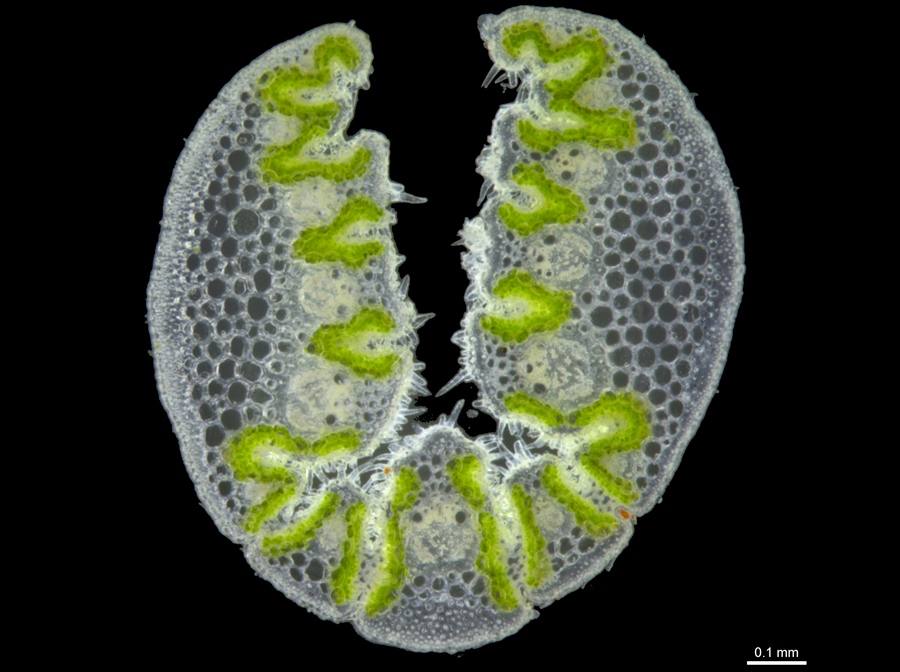
T. veniciae leaf section.

T. veniciae spikelet (longer extreme).

T. veniciae glumes.

T. veniciae lemmas.

T. veniciae lemma base.

T. veniciae palea.
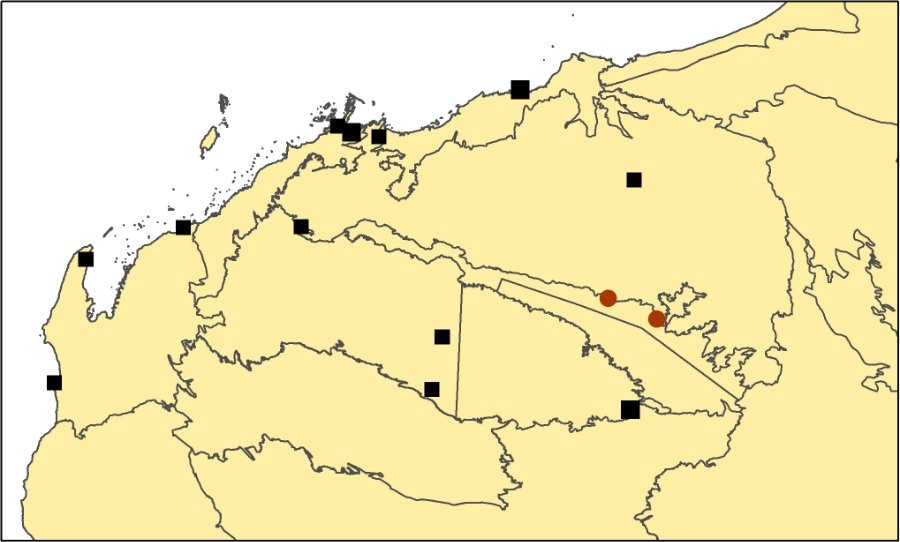
T. veniciae map.
Name
Triodia veniciae M.D.Barrett, ined.
Citation
Nuytsia, in press, (2017)
Derivation
veniciae — after Venicia De San Miguel (née Giannasi), from Roy Hill Environment Department, who was instrumental in the discovery, collection of type material and surveys for this species.Common name
Roy Hill Spinifex
Synonyms
None
Diagnostic features
Foliage strongly resinous; leaves epistomatous (soft-type); leaf sheaths with sparse to moderately dense hairs on surfaces; longest inflorescence branches with 7–14 spikelets; lower glume 5.8–8.5 mm long, narrowly lanceolate, L:W 5–6.8; lemmas lobed, the lobes narrow and, sometimes with the appearance of short awns; lemma body uniformly textured, with hairs arranged in longitudinal rows; lowest lemma midlobe 2.1–3.2 mm long; on shale slopes; only in Chichester Range north of Fortescue River valley and east of the Great Northern Highway.
Habitat
Occurs on shale slopes and tops of low hills. Apparently absent from ironstone mesas and plateaus, which form the dominant rocky habitat in the south-eastern Chichester range.
Distribution and frequency
Endemic to the Pilbara region. Only known from a few populations in the eastern Chichester Range on the north side of the Fortescue Valley. Populations are scattered in the landscape, and are much less frequent than Triodia epactia, which is widespread in the area.
Similar species
Triodia veniciae belongs to the Soft group, sharing the epistomatous (soft-type) leaf blades. Amongst the Soft group species, T. veniciae and T. karijini are the only species sharing the combination of resinous foliage, hairy sheath surfaces, narrowly lanceolate glumes (L:W>4) and uniformly textured (not bitextured) lemmas — both species are also unusual in having acuminate to sub-awned lemma lobes, intermediate between the prominent awns or clearly un-awned condition of other species.
Triodia karijini is the most similar species morphologically, sharing the narrowly triangular to sub-awned lemmas lobes 2–3.2 mm long and hairy leaf sheath surfaces. Triodia veniciae is copiously resinous (non-resinous to weakly resinous in T. karijini), and palea with hairs restricted to central zone (palea glabrous in T. karijini), and 7–14 spikelets on longest basal panicle branches (3–6 spikelets in T. karijini). Triodia karijini is only known from above 900 m on ironstone ridges on mountains in the Hamersley Range, while T. veniciae is only known from shale slopes in the Chichester Range north of the Fortescue River.
Triodia veniciae is also similar to T. melvillei, sharing very resinous habit, narrow glumes and lemma hairs concentrated in longitudinal rows. However, the lemmas are 3-awned in T. melvillei (lowest lemma midlobe 5.5–6.8 mm long in Pilbara specimens), compared to lobed to sub-awned with lowest lemma midlobe 2.1–3.2 mm long in T. veniciae. The leaf sheaths are hairy on their surfaces in T. veniciae, whereas they are glabrous in T. melvillei. The paleas in T. veniciae have hairs restricted to the central area, whereas paleas of T. melvillei are either glabrous or have scattered hairs over the lower 1/2–3/4. Triodia veniciae is restricted to the Chichester Range north of the Fortescue River, while T. melvillei occurs in the Pilbara south of the Fortescue River, south and east into central Australia.
Conservation status
Priority One recommended.
Identification without florets
In the south-eastern Chichester sub-region, the only species with either resinous foliage or epistomatous (soft-type) leaves are T. epactia and T. veniciae. Triodia veniciae and T. epactia co-occur, and can be distinguished by leaf sheath surfaces (hairy in T. veniciae, not hairy in T. epactia) and glume shape (L:W4 in T. epactia, L:W≥5 in T. veniciae).
Triodia schinzii also occurs in the eastern Chichester sub-region, but can be distinguished by lacking hairs on the leaf sheaths (hairy in T. veniciae), having glumes >12 mm long (5.8–8.5 mm long in T. veniciae), and habit on sand or sandy loam flats rather than rocky slopes.
Triodia basitricha also occurs in the eastern Chichester sub-region, and shares hairy sheath surfaces and habit with T. veniciae. Triodia basitricha has longer lower glumes 10–12.5 mm long (5.4–8.5 mm long in T. veniciae) and 4–5 spikelets on longest basal branches (7–14 in T. veniciae).
Variation
A uniform species as currently known.
Notes
Triodia veniciae was not represented in Lazarides (1997), Lazarides et al. (2005) or Ausgrass (Sharp & Simon, 2002; Simon & Alonso, 2014).
A full description of T. veniciae can be found in Barrett (2017a).